แบคทีเรียมี DNA หรือไม่? สารพันธุกรรมของแบคทีเรีย ดีเอ็นเอของแบคทีเรีย ดีเอ็นเอของแบคทีเรียในสเมียร์
แบคทีเรียเป็นกลุ่มสิ่งมีชีวิตที่เก่าแก่ที่สุดที่มีอยู่บนโลกในปัจจุบัน แบคทีเรียกลุ่มแรกอาจปรากฏขึ้นเมื่อกว่า 3.5 พันล้านปีก่อน และเป็นเวลาเกือบพันล้านปีแล้วที่พวกมันเป็นสิ่งมีชีวิตเพียงชนิดเดียวในโลกของเรา เนื่องจากสิ่งเหล่านี้เป็นตัวแทนแรกของธรรมชาติที่มีชีวิต ร่างกายของพวกเขาจึงมีโครงสร้างดั้งเดิม
เมื่อเวลาผ่านไป โครงสร้างของพวกเขามีความซับซ้อนมากขึ้น แต่จนถึงทุกวันนี้ แบคทีเรียถือเป็นสิ่งมีชีวิตเซลล์เดียวดึกดำบรรพ์ที่สุด เป็นที่น่าสนใจที่แบคทีเรียบางชนิดยังคงรักษาลักษณะดั้งเดิมของบรรพบุรุษโบราณเอาไว้ พบได้ในแบคทีเรียที่อาศัยอยู่ในบ่อน้ำพุร้อนกำมะถันและโคลนที่เป็นพิษที่ด้านล่างของอ่างเก็บน้ำ
แบคทีเรียส่วนใหญ่ไม่มีสี มีเพียงไม่กี่สีม่วงหรือสีเขียว แต่อาณานิคมของแบคทีเรียหลายชนิดมีสีสดใสซึ่งเกิดจากการปล่อยสารที่มีสีออกสู่สิ่งแวดล้อมหรือการสร้างเม็ดสีของเซลล์
ผู้ค้นพบโลกแห่งแบคทีเรียคือ Antony Leeuwenhoek นักธรรมชาติวิทยาชาวดัตช์แห่งศตวรรษที่ 17 ผู้สร้างกล้องจุลทรรศน์ขยายภาพที่สมบูรณ์แบบเป็นครั้งแรกซึ่งสามารถขยายวัตถุได้ 160-270 เท่า
แบคทีเรียจัดอยู่ในประเภทโปรคาริโอตและแบ่งออกเป็นอาณาจักรที่แยกจากกัน - แบคทีเรีย
รูปร่าง
แบคทีเรียเป็นสิ่งมีชีวิตมากมายและหลากหลาย มีรูปร่างแตกต่างกันไป
| ชื่อของแบคทีเรีย | รูปร่างของแบคทีเรีย | ภาพแบคทีเรีย |
| ค็อกซี่ | มีลักษณะเป็นลูกบอล | |
| บาซิลลัส |  | มีลักษณะเป็นแท่ง |
| วิบริโอ | รูปทรงจุลภาค | |
| สไปริลลัม |  | เกลียว |
| สเตรปโตคอคกี้ |  | สายโซ่ค็อกกี้ |
| สแตฟิโลคอคคัส |  | กลุ่มของ cocci |
| ดิพโลคอคคัส | แบคทีเรียทรงกลม 2 ตัวอยู่ในแคปซูลเมือกเดียว |
วิธีการขนส่ง
ในบรรดาแบคทีเรียนั้นมีรูปแบบเคลื่อนที่และไม่เคลื่อนที่ การเคลื่อนไหวเคลื่อนที่เนื่องจากการหดตัวคล้ายคลื่นหรือด้วยความช่วยเหลือของแฟลเจลลา (เกลียวเกลียวที่บิดเป็นเกลียว) ซึ่งประกอบด้วยโปรตีนพิเศษที่เรียกว่าแฟลเจลลิน อาจมีแฟลเจลลาอย่างน้อยหนึ่งรายการ ในแบคทีเรียบางชนิด พวกมันจะอยู่ที่ปลายด้านหนึ่งของเซลล์ ส่วนบางชนิดจะอยู่ที่ 2 อันหรือทั่วพื้นผิวทั้งหมด

แต่การเคลื่อนไหวก็มีอยู่ในแบคทีเรียอื่น ๆ อีกมากมายที่ไม่มีแฟลเจลลา ดังนั้นแบคทีเรียที่ปกคลุมด้านนอกด้วยเมือกจึงสามารถเคลื่อนไหวได้
แบคทีเรียในน้ำและในดินบางชนิดที่ไม่มีแฟลเจลลาจะมีแวคิวโอลของก๊าซอยู่ในไซโตพลาสซึม อาจมีแวคิวโอล 40-60 ในเซลล์ แต่ละคนเต็มไปด้วยก๊าซ (น่าจะเป็นไนโตรเจน) ด้วยการควบคุมปริมาณก๊าซในแวคิวโอล แบคทีเรียในน้ำสามารถจมลงในคอลัมน์น้ำหรือลอยขึ้นสู่ผิวน้ำได้ และแบคทีเรียในดินสามารถเคลื่อนที่ในเส้นเลือดฝอยในดินได้
ที่อยู่อาศัย
เนื่องจากความเรียบง่ายของการจัดระเบียบและไม่โอ้อวด แบคทีเรียจึงแพร่หลายในธรรมชาติ แบคทีเรียพบได้ทุกที่: ในหยดน้ำแม้แต่น้ำพุที่บริสุทธิ์ที่สุด ในเมล็ดดิน ในอากาศ บนโขดหิน ในหิมะขั้วโลก ทรายทะเลทราย บนพื้นมหาสมุทร ในน้ำมันที่สกัดจากส่วนลึกที่ยิ่งใหญ่ และแม้แต่ใน น้ำพุร้อนที่มีอุณหภูมิประมาณ 80 องศาเซลเซียส อาศัยอยู่บนพืช ผลไม้ สัตว์ต่างๆ และในมนุษย์ในลำไส้ ช่องปาก แขนขา และบนพื้นผิวของร่างกาย
แบคทีเรียเป็นสิ่งมีชีวิตที่เล็กที่สุดและมีจำนวนมากที่สุด เนื่องจากมีขนาดเล็ก จึงเจาะเข้าไปในรอยแตก รอยแยก หรือรูพรุนต่างๆ ได้อย่างง่ายดาย แข็งแกร่งมากและปรับให้เข้ากับสภาพความเป็นอยู่ต่างๆ พวกเขาทนต่อการแห้ง ความเย็นจัด และความร้อนสูงถึง 90°C โดยไม่สูญเสียความสามารถในการอยู่รอด
ในทางปฏิบัติแล้วไม่มีสถานที่ใดบนโลกที่ไม่พบแบคทีเรีย แต่จะมีปริมาณที่แตกต่างกันออกไป สภาพความเป็นอยู่ของแบคทีเรียนั้นแตกต่างกันไป บางชนิดต้องการออกซิเจนในบรรยากาศ บางชนิดไม่ต้องการและสามารถอาศัยอยู่ในสภาพแวดล้อมที่ปราศจากออกซิเจนได้
ในอากาศ: แบคทีเรียลอยขึ้นสู่บรรยากาศชั้นบนได้ไกลถึง 30 กม. และอื่น ๆ.
มีจำนวนมากโดยเฉพาะในดิน ดิน 1 กรัม มีแบคทีเรียนับร้อยล้านตัว
ในน้ำ: ในชั้นผิวน้ำในอ่างเก็บน้ำเปิด แบคทีเรียในน้ำที่เป็นประโยชน์จะดูดซับสารอินทรีย์ที่ตกค้าง
ในสิ่งมีชีวิต: แบคทีเรียที่ทำให้เกิดโรคเข้าสู่ร่างกายจากสภาพแวดล้อมภายนอก แต่ภายใต้สภาวะที่เอื้ออำนวยเท่านั้นที่ทำให้เกิดโรค ซิมไบโอติกอาศัยอยู่ในอวัยวะย่อยอาหาร ช่วยย่อยและดูดซึมอาหาร และสังเคราะห์วิตามิน
โครงสร้างภายนอก
เซลล์แบคทีเรียถูกปกคลุมไปด้วยเปลือกที่มีความหนาแน่นเป็นพิเศษ - ผนังเซลล์ซึ่งทำหน้าที่ป้องกันและรองรับและยังทำให้แบคทีเรียมีรูปร่างที่มีลักษณะเฉพาะถาวรอีกด้วย ผนังเซลล์ของแบคทีเรียมีลักษณะคล้ายกับผนังเซลล์พืช สามารถซึมผ่านได้: สารอาหารจะผ่านเข้าไปในเซลล์ได้อย่างอิสระและผลิตภัณฑ์เมตาบอลิซึมจะออกสู่สิ่งแวดล้อม บ่อยครั้งที่แบคทีเรียสร้างชั้นป้องกันเมือกเพิ่มเติมที่ด้านบนของผนังเซลล์ซึ่งก็คือแคปซูล ความหนาของแคปซูลอาจมากกว่าเส้นผ่านศูนย์กลางของเซลล์หลายเท่า แต่ก็อาจมีขนาดเล็กมากได้เช่นกัน แคปซูลไม่ใช่ส่วนสำคัญของเซลล์แต่ถูกสร้างขึ้นขึ้นอยู่กับสภาวะที่แบคทีเรียพบตัวเอง ช่วยปกป้องแบคทีเรียไม่ให้แห้ง
บนพื้นผิวของแบคทีเรียบางชนิดจะมีแฟลเจลลายาว (หนึ่ง สอง หรือมาก) หรือวิลลี่บางสั้น ความยาวของแฟลเจลลาอาจมากกว่าขนาดลำตัวของแบคทีเรียหลายเท่า แบคทีเรียเคลื่อนที่ด้วยความช่วยเหลือของแฟลเจลลาและวิลลี่

โครงสร้างภายใน
ภายในเซลล์แบคทีเรียจะมีไซโตพลาสซึมหนาแน่นและเคลื่อนที่ไม่ได้ มีโครงสร้างเป็นชั้นไม่มีแวคิวโอลดังนั้นโปรตีน (เอนไซม์) และสารอาหารสำรองต่าง ๆ จึงอยู่ในสารของไซโตพลาสซึมนั่นเอง เซลล์แบคทีเรียไม่มีนิวเคลียส สารที่มีข้อมูลทางพันธุกรรมจะกระจุกตัวอยู่ที่ส่วนกลางของเซลล์ แบคทีเรีย - กรดนิวคลีอิก - ดีเอ็นเอ แต่สารนี้ไม่ได้ก่อตัวเป็นนิวเคลียส
โครงสร้างภายในของเซลล์แบคทีเรียมีความซับซ้อนและมีลักษณะเฉพาะของตัวเอง ไซโตพลาสซึมถูกแยกออกจากผนังเซลล์โดยเยื่อหุ้มเซลล์ ในไซโตพลาสซึมมีสารหลักหรือเมทริกซ์ไรโบโซมและโครงสร้างเมมเบรนจำนวนเล็กน้อยที่ทำหน้าที่หลายอย่าง (อะนาล็อกของไมโตคอนเดรีย, เรติเคิลเอนโดพลาสมิก, อุปกรณ์ Golgi) พลาสซึมของเซลล์แบคทีเรียมักประกอบด้วยเม็ดที่มีรูปร่างและขนาดต่างๆ เม็ดอาจประกอบด้วยสารประกอบที่ทำหน้าที่เป็นแหล่งพลังงานและคาร์บอน พบหยดไขมันในเซลล์แบคทีเรียด้วย
ในส่วนกลางของเซลล์สารนิวเคลียร์จะถูกแปลเป็นภาษาท้องถิ่น - DNA ซึ่งไม่ได้ถูกคั่นด้วยไซโตพลาสซึมด้วยเมมเบรน นี่คืออะนาล็อกของนิวเคลียส - นิวเคลียส นิวคลอยด์ไม่มีเมมเบรน นิวคลีโอลัส หรือชุดโครโมโซม
วิธีการรับประทาน
แบคทีเรียมีวิธีให้อาหารที่แตกต่างกัน ในหมู่พวกเขามีออโตโทรฟและเฮเทอโรโทรฟ ออโตโทรฟเป็นสิ่งมีชีวิตที่สามารถผลิตสารอินทรีย์เพื่อเป็นสารอาหารได้อย่างอิสระ
พืชต้องการไนโตรเจน แต่ไม่สามารถดูดซับไนโตรเจนจากอากาศได้เอง แบคทีเรียบางชนิดรวมโมเลกุลไนโตรเจนในอากาศเข้ากับโมเลกุลอื่น ๆ ส่งผลให้เกิดสารที่มีอยู่ในพืช
แบคทีเรียเหล่านี้สะสมอยู่ในเซลล์ของรากอ่อน ซึ่งนำไปสู่การก่อตัวของรากที่หนาขึ้น เรียกว่าก้อนเนื้อ ก้อนดังกล่าวก่อตัวบนรากของพืชตระกูลถั่วและพืชอื่น ๆ
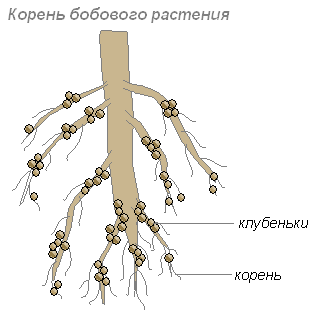
รากให้คาร์โบไฮเดรตแก่แบคทีเรีย และแบคทีเรียให้สารที่มีไนโตรเจนแก่รากซึ่งพืชสามารถดูดซึมได้ การอยู่ร่วมกันของพวกเขาเป็นประโยชน์ร่วมกัน
รากพืชจะหลั่งสารอินทรีย์จำนวนมาก (น้ำตาล กรดอะมิโน และอื่นๆ) ที่แบคทีเรียกินเข้าไป ดังนั้นโดยเฉพาะแบคทีเรียจำนวนมากจึงเกาะตัวอยู่ในชั้นดินที่อยู่รอบราก แบคทีเรียเหล่านี้จะเปลี่ยนเศษซากพืชที่ตายแล้วให้เป็นสารที่มีอยู่ในพืช ชั้นดินนี้เรียกว่าไรโซสเฟียร์
มีสมมติฐานหลายประการเกี่ยวกับการแทรกซึมของแบคทีเรียปมเข้าไปในเนื้อเยื่อราก:
- ผ่านความเสียหายต่อเนื้อเยื่อผิวหนังชั้นนอกและเยื่อหุ้มสมอง
- ผ่านขนราก
- ผ่านเยื่อหุ้มเซลล์อ่อนเท่านั้น
- ขอบคุณแบคทีเรียสหายที่ผลิตเอนไซม์เพคติโนไลติก
- เนื่องจากการกระตุ้นการสังเคราะห์กรด B-indoleacetic จากทริปโตเฟน ซึ่งมักพบอยู่ในสารคัดหลั่งของรากพืช
กระบวนการนำแบคทีเรียปมเข้าไปในเนื้อเยื่อรากประกอบด้วยสองขั้นตอน:
- การติดเชื้อของขนราก
- กระบวนการสร้างปม
ในกรณีส่วนใหญ่ เซลล์ที่บุกรุกจะทวีคูณอย่างแข็งขัน ก่อให้เกิดสิ่งที่เรียกว่าเส้นติดเชื้อ และในรูปแบบของเส้นดังกล่าว จะเคลื่อนเข้าสู่เนื้อเยื่อพืช แบคทีเรียที่เป็นปมที่โผล่ออกมาจากด้ายที่ติดเชื้อจะยังคงเพิ่มจำนวนในเนื้อเยื่อของโฮสต์

เซลล์พืชที่เต็มไปด้วยเซลล์ของแบคทีเรียที่มีการขยายตัวอย่างรวดเร็วจะเริ่มแบ่งตัวอย่างรวดเร็ว การเชื่อมต่อของปมอ่อนกับรากของพืชตระกูลถั่วนั้นเกิดจากการรวมกลุ่มของเส้นใยหลอดเลือด ในระหว่างการทำงาน ก้อนเนื้อมักจะหนาแน่น เมื่อถึงเวลาที่มีกิจกรรมที่เหมาะสมที่สุด ก้อนจะกลายเป็นสีชมพู (ต้องขอบคุณเม็ดสีเลฮีโมโกลบิน) เฉพาะแบคทีเรียที่มีเลฮีโมโกลบินเท่านั้นที่สามารถตรึงไนโตรเจนได้
แบคทีเรียที่เป็นก้อนกลมสร้างปุ๋ยไนโตรเจนได้หลายสิบหลายร้อยกิโลกรัมต่อเฮกตาร์ของดิน
การเผาผลาญอาหาร
แบคทีเรียแตกต่างกันในการเผาผลาญ ในบางกรณีมันเกิดขึ้นพร้อมกับการมีส่วนร่วมของออกซิเจน บางอย่างเกิดขึ้นโดยไม่มีมัน
แบคทีเรียส่วนใหญ่กินสารอินทรีย์สำเร็จรูป มีเพียงไม่กี่ชนิดเท่านั้น (สีน้ำเงินเขียวหรือไซยาโนแบคทีเรีย) ที่สามารถสร้างสารอินทรีย์จากอนินทรีย์ได้ มีบทบาทสำคัญในการสะสมของออกซิเจนในชั้นบรรยากาศของโลก
แบคทีเรียดูดซับสารจากภายนอก ฉีกโมเลกุลออกเป็นชิ้น ๆ ประกอบเปลือกของพวกมันจากส่วนเหล่านี้และเติมเต็มเนื้อหา (นี่คือวิธีที่พวกมันเติบโต) และโยนโมเลกุลที่ไม่จำเป็นออกไป เปลือกและเยื่อหุ้มของแบคทีเรียช่วยให้สามารถดูดซับเฉพาะสารที่จำเป็นเท่านั้น

หากเปลือกและเยื่อหุ้มของแบคทีเรียไม่สามารถซึมผ่านได้อย่างสมบูรณ์ ก็จะไม่มีสารใดเข้าไปในเซลล์ได้ หากสารเหล่านั้นซึมผ่านได้ สารในเซลล์ก็จะผสมกับตัวกลางซึ่งเป็นสารละลายที่แบคทีเรียอาศัยอยู่ เพื่อความอยู่รอด แบคทีเรียจำเป็นต้องมีเปลือกที่ช่วยให้สารที่จำเป็นสามารถผ่านไปได้ แต่ไม่ใช่สารที่ไม่จำเป็น
แบคทีเรียดูดซับสารอาหารที่อยู่ใกล้มัน จะเกิดอะไรขึ้นต่อไป? หากสามารถเคลื่อนที่ได้อย่างอิสระ (โดยการขยับแฟลเจลลัมหรือดันเมือกกลับ) ก็จะเคลื่อนที่จนกว่าจะพบสารที่จำเป็น

ถ้ามันไม่สามารถเคลื่อนที่ได้ มันก็จะรอจนกระทั่งการแพร่กระจาย (ความสามารถของโมเลกุลของสารหนึ่งในการแทรกซึมเข้าไปในกลุ่มโมเลกุลของสารอื่น) จะนำโมเลกุลที่จำเป็นเข้าไป
แบคทีเรียร่วมกับจุลินทรีย์กลุ่มอื่นทำหน้าที่ทางเคมีจำนวนมหาศาล โดยการแปลงสารประกอบต่างๆ พวกมันจะได้รับพลังงานและสารอาหารที่จำเป็นต่อชีวิต กระบวนการเมตาบอลิซึม วิธีการรับพลังงาน และความต้องการวัสดุในการสร้างสารในร่างกายมีความหลากหลายในแบคทีเรีย
แบคทีเรียชนิดอื่นๆ ตอบสนองความต้องการทั้งหมดสำหรับคาร์บอนที่จำเป็นสำหรับการสังเคราะห์สารอินทรีย์ในร่างกาย โดยแทนที่สารประกอบอนินทรีย์ พวกมันถูกเรียกว่าออโตโทรฟ แบคทีเรียออโตโทรฟิกสามารถสังเคราะห์สารอินทรีย์จากสารอนินทรีย์ได้ ในหมู่พวกเขาคือ:
การสังเคราะห์ทางเคมี
การใช้พลังงานรังสีเป็นสิ่งสำคัญที่สุด แต่ไม่ใช่วิธีเดียวที่จะสร้างอินทรียวัตถุจากคาร์บอนไดออกไซด์และน้ำ เป็นที่รู้กันว่าแบคทีเรียไม่ได้ใช้แสงแดดเป็นแหล่งพลังงานในการสังเคราะห์ แต่เป็นพลังงานของพันธะเคมีที่เกิดขึ้นในเซลล์ของสิ่งมีชีวิตระหว่างการออกซิเดชันของสารประกอบอนินทรีย์บางชนิด ได้แก่ ไฮโดรเจนซัลไฟด์ ซัลเฟอร์ แอมโมเนีย ไฮโดรเจน กรดไนตริก สารประกอบเหล็กของ เหล็กและแมงกานีส พวกเขาใช้อินทรียวัตถุที่เกิดจากพลังงานเคมีนี้เพื่อสร้างเซลล์ในร่างกาย ดังนั้นกระบวนการนี้เรียกว่าการสังเคราะห์ทางเคมี
กลุ่มจุลินทรีย์สังเคราะห์ทางเคมีที่สำคัญที่สุดคือแบคทีเรียไนตริไฟอิง แบคทีเรียเหล่านี้อาศัยอยู่ในดินและออกซิไดซ์แอมโมเนียที่เกิดขึ้นระหว่างการสลายตัวของสารอินทรีย์ตกค้างเป็นกรดไนตริก หลังทำปฏิกิริยากับสารประกอบแร่ในดินกลายเป็นเกลือของกรดไนตริก กระบวนการนี้เกิดขึ้นในสองขั้นตอน
แบคทีเรียเหล็กเปลี่ยนเหล็กเป็นเหล็กออกไซด์ เหล็กไฮดรอกไซด์ที่เกิดขึ้นจะเกาะตัวและก่อตัวเป็นแร่เหล็กบึง
จุลินทรีย์บางชนิดมีอยู่เนื่องจากการออกซิเดชันของโมเลกุลไฮโดรเจน ดังนั้นจึงให้วิธีการทางโภชนาการแบบออโตโทรฟิค
คุณลักษณะเฉพาะของแบคทีเรียไฮโดรเจนคือความสามารถในการเปลี่ยนไปสู่วิถีชีวิตแบบเฮเทอโรโทรฟิคเมื่อได้รับสารประกอบอินทรีย์และไม่มีไฮโดรเจน
ดังนั้น chemoautotrophs จึงเป็นออโตโทรฟทั่วไปเนื่องจากพวกมันสังเคราะห์สารประกอบอินทรีย์ที่จำเป็นจากสารอนินทรีย์อย่างอิสระและไม่ได้นำพวกมันสำเร็จรูปจากสิ่งมีชีวิตอื่นเช่นเฮเทอโรโทรฟ แบคทีเรียเคมีบำบัดแตกต่างจากพืชโฟโตโทรฟิคตรงที่พวกมันเป็นอิสระจากแสงเป็นแหล่งพลังงาน
การสังเคราะห์ด้วยแสงของแบคทีเรีย
แบคทีเรียกำมะถันที่มีเม็ดสีบางชนิด (สีม่วง, สีเขียว) ซึ่งมีเม็ดสีเฉพาะ - แบคทีเรียคลอโรฟิลล์สามารถดูดซับพลังงานแสงอาทิตย์ได้ด้วยความช่วยเหลือซึ่งทำให้ไฮโดรเจนซัลไฟด์ในร่างกายถูกทำลายและปล่อยอะตอมไฮโดรเจนเพื่อฟื้นฟูสารประกอบที่เกี่ยวข้อง กระบวนการนี้มีอะไรเหมือนกันมากกับการสังเคราะห์ด้วยแสง และแตกต่างตรงที่ในแบคทีเรียสีม่วงและสีเขียว ผู้บริจาคไฮโดรเจนคือไฮโดรเจนซัลไฟด์ (บางครั้งก็เป็นกรดคาร์บอกซิลิก) และในพืชสีเขียวก็คือน้ำ ในทั้งสองอย่างการแยกและการถ่ายโอนไฮโดรเจนเกิดขึ้นเนื่องจากพลังงานของรังสีดวงอาทิตย์ที่ถูกดูดซับ
การสังเคราะห์ด้วยแสงของแบคทีเรียซึ่งเกิดขึ้นโดยไม่มีการปล่อยออกซิเจนเรียกว่าการลดแสง การลดคาร์บอนไดออกไซด์ด้วยแสงนั้นสัมพันธ์กับการถ่ายโอนไฮโดรเจนไม่ใช่จากน้ำ แต่จากไฮโดรเจนซัลไฟด์:
6СО 2 +12Н 2 S+hv → С6Н 12 О 6 +12S=6Н 2 О
ความสำคัญทางชีวภาพของการสังเคราะห์ทางเคมีและการสังเคราะห์ด้วยแสงของแบคทีเรียในระดับดาวเคราะห์นั้นค่อนข้างเล็ก มีเพียงแบคทีเรียสังเคราะห์ทางเคมีเท่านั้นที่มีบทบาทสำคัญในกระบวนการหมุนเวียนของกำมะถันในธรรมชาติ ซัลเฟอร์จะถูกดูดซึมโดยพืชสีเขียวในรูปของเกลือของกรดซัลฟิวริกและกลายเป็นส่วนหนึ่งของโมเลกุลโปรตีน นอกจากนี้ เมื่อซากพืชและสัตว์ที่ตายแล้วถูกทำลายโดยแบคทีเรียที่เน่าเปื่อย ซัลเฟอร์จะถูกปล่อยออกมาในรูปของไฮโดรเจนซัลไฟด์ ซึ่งถูกออกซิไดซ์โดยแบคทีเรียซัลเฟอร์จนกลายเป็นกำมะถันอิสระ (หรือกรดซัลฟิวริก) ทำให้เกิดซัลไฟต์ในดินที่พืชสามารถเข้าถึงได้ แบคทีเรียเคมีบำบัดและโฟโตออโตโทรฟิคมีความสำคัญในวงจรไนโตรเจนและซัลเฟอร์
การสร้างสปอร์
สปอร์ก่อตัวภายในเซลล์แบคทีเรีย ในระหว่างกระบวนการสร้างสปอร์ เซลล์แบคทีเรียจะผ่านกระบวนการทางชีวเคมีหลายอย่าง ปริมาณน้ำอิสระในนั้นลดลงและกิจกรรมของเอนไซม์ลดลง สิ่งนี้ทำให้มั่นใจได้ถึงความต้านทานของสปอร์ต่อสภาพแวดล้อมที่ไม่เอื้ออำนวย (อุณหภูมิสูง ความเข้มข้นของเกลือสูง การแห้ง ฯลฯ) การสร้างสปอร์เป็นลักษณะของแบคทีเรียกลุ่มเล็กๆ เท่านั้น

สปอร์เป็นขั้นตอนเสริมในวงจรชีวิตของแบคทีเรีย การสร้างสปอร์เริ่มต้นจากการขาดสารอาหารหรือการสะสมของผลิตภัณฑ์จากการเผาผลาญเท่านั้น แบคทีเรียในรูปสปอร์สามารถคงอยู่เฉยๆได้เป็นเวลานาน สปอร์ของแบคทีเรียสามารถทนต่อการเดือดเป็นเวลานานและการแช่แข็งที่ยาวนานมาก เมื่อสภาวะเอื้ออำนวยสปอร์จะงอกและมีชีวิตได้ สปอร์ของแบคทีเรียเป็นการปรับตัวเพื่อความอยู่รอดในสภาวะที่ไม่เอื้ออำนวย
การสืบพันธุ์
แบคทีเรียสืบพันธุ์โดยการแบ่งเซลล์หนึ่งออกเป็นสองเซลล์ เมื่อถึงขนาดที่กำหนด แบคทีเรียจะแบ่งออกเป็นแบคทีเรียที่เหมือนกันสองตัว จากนั้นพวกมันแต่ละตัวก็เริ่มกินอาหาร เติบโต แบ่งตัว และอื่นๆ

หลังจากการยืดตัวของเซลล์ กะบังตามขวางจะค่อยๆ ก่อตัวขึ้น จากนั้นเซลล์ลูกสาวจะแยกออกจากกัน ในแบคทีเรียหลายชนิด หลังจากแบ่งเซลล์แล้ว ภายใต้เงื่อนไขบางประการ เซลล์ยังคงเชื่อมต่อกันเป็นกลุ่มลักษณะเฉพาะ ในกรณีนี้ ขึ้นอยู่กับทิศทางของระนาบการแบ่งและจำนวนการแบ่ง รูปร่างที่แตกต่างกันจะเกิดขึ้น การสืบพันธุ์โดยการแตกหน่อเกิดขึ้นเป็นข้อยกเว้นในแบคทีเรีย
ภายใต้สภาวะที่เอื้ออำนวย การแบ่งเซลล์ในแบคทีเรียจำนวนมากจะเกิดขึ้นทุกๆ 20-30 นาที ด้วยการแพร่พันธุ์อย่างรวดเร็วเช่นนี้ ลูกของแบคทีเรียหนึ่งตัวใน 5 วันจะสามารถสร้างมวลที่เต็มทะเลและมหาสมุทรได้ การคำนวณอย่างง่ายแสดงให้เห็นว่าสามารถเกิดขึ้นได้ 72 รุ่น (720,000,000,000,000,000,000 เซลล์) ต่อวัน หากแปลงเป็นน้ำหนัก - 4720 ตัน อย่างไรก็ตาม สิ่งนี้ไม่ได้เกิดขึ้นในธรรมชาติ เนื่องจากแบคทีเรียส่วนใหญ่ตายอย่างรวดเร็วภายใต้อิทธิพลของแสงแดด ทำให้แห้ง ขาดอาหาร อุณหภูมิถึง 65-100°C ซึ่งเป็นผลมาจากการต่อสู้ระหว่างสายพันธุ์ เป็นต้น
แบคทีเรีย (1) เมื่อดูดซึมอาหารได้เพียงพอ จะเพิ่มขนาด (2) และเริ่มเตรียมการสืบพันธุ์ (การแบ่งเซลล์) DNA ของมัน (ในแบคทีเรีย โมเลกุล DNA จะถูกปิดอยู่ในวงแหวน) จะเพิ่มเป็นสองเท่า (แบคทีเรียสร้างสำเนาของโมเลกุลนี้) โมเลกุล DNA ทั้งสอง (3,4) พบว่าตัวเองติดอยู่กับผนังของแบคทีเรีย และเมื่อแบคทีเรียยืดออก ก็จะแยกออกจากกัน (5,6) ขั้นแรกนิวคลีโอไทด์จะแบ่งตัว จากนั้นจึงเกิดไซโตพลาสซึม

หลังจากการแยกโมเลกุล DNA ทั้งสองออกจากกัน เกิดการหดตัวบนแบคทีเรีย ซึ่งค่อยๆ แบ่งร่างกายของแบคทีเรียออกเป็นสองส่วน โดยแต่ละส่วนจะมีโมเลกุล DNA (7)
มันเกิดขึ้น (ใน Bacillus subtilis) ที่แบคทีเรียสองตัวเกาะติดกันและมีสะพานเชื่อมระหว่างพวกมัน (1,2)

จัมเปอร์ขนส่ง DNA จากแบคทีเรียหนึ่งไปยังอีกแบคทีเรียหนึ่ง (3) เมื่ออยู่ในแบคทีเรียตัวเดียว โมเลกุล DNA จะพันกัน และเกาะติดกันในบางแห่ง (4) จากนั้นจึงแลกเปลี่ยนส่วนต่างๆ (5)
บทบาทของแบคทีเรียในธรรมชาติ
ไกร์
แบคทีเรียเป็นตัวเชื่อมโยงที่สำคัญที่สุดในวัฏจักรทั่วไปของสารในธรรมชาติ พืชสร้างสารอินทรีย์ที่ซับซ้อนจากคาร์บอนไดออกไซด์ น้ำ และเกลือแร่ในดิน สารเหล่านี้กลับคืนสู่ดินพร้อมกับเชื้อราที่ตายแล้ว พืช และซากสัตว์ แบคทีเรียจะสลายสารที่ซับซ้อนให้กลายเป็นสารง่ายๆ จากนั้นพืชจึงนำไปใช้
แบคทีเรียทำลายสารอินทรีย์ที่ซับซ้อนของพืชที่ตายแล้วและซากสัตว์ การขับถ่ายของสิ่งมีชีวิตและของเสียต่างๆ แบคทีเรีย saprophytic ที่สลายตัวโดยการกินสารอินทรีย์เหล่านี้จะเปลี่ยนพวกมันให้กลายเป็นฮิวมัส สิ่งเหล่านี้เป็นระเบียบเรียบร้อยของโลกของเรา ดังนั้นแบคทีเรียจึงมีส่วนร่วมในวงจรของสารในธรรมชาติ
การก่อตัวของดิน
เนื่องจากแบคทีเรียกระจายตัวไปเกือบทุกที่และเกิดขึ้นเป็นจำนวนมาก แบคทีเรียจึงกำหนดกระบวนการต่างๆ ที่เกิดขึ้นในธรรมชาติเป็นส่วนใหญ่ ในฤดูใบไม้ร่วงใบไม้ของต้นไม้และพุ่มไม้ร่วงหล่นยอดหญ้าเหนือพื้นดินตายกิ่งเก่าร่วงหล่นและลำต้นของต้นไม้เก่าก็ร่วงหล่นเป็นครั้งคราว ทั้งหมดนี้ค่อยๆ กลายเป็นฮิวมัส ใน 1 ซม.3 ชั้นผิวของดินป่าประกอบด้วยแบคทีเรียในดิน saprophytic หลายร้อยล้านชนิด แบคทีเรียเหล่านี้เปลี่ยนฮิวมัสให้เป็นแร่ธาตุต่างๆ ที่สามารถดูดซึมได้จากดินโดยรากพืช
แบคทีเรียในดินบางชนิดสามารถดูดซับไนโตรเจนจากอากาศเพื่อใช้ในกระบวนการสำคัญได้ แบคทีเรียตรึงไนโตรเจนเหล่านี้อาศัยอยู่อย่างอิสระหรืออาศัยอยู่ที่รากของพืชตระกูลถั่ว เมื่อเจาะรากของพืชตระกูลถั่วแล้วแบคทีเรียเหล่านี้ทำให้เกิดการเจริญเติบโตของเซลล์รากและการก่อตัวของก้อนบนพวกมัน
แบคทีเรียเหล่านี้ผลิตสารประกอบไนโตรเจนที่พืชใช้ แบคทีเรียได้รับคาร์โบไฮเดรตและเกลือแร่จากพืช ดังนั้นจึงมีความสัมพันธ์ใกล้ชิดระหว่างพืชตระกูลถั่วกับแบคทีเรียที่เป็นปมซึ่งเป็นประโยชน์ต่อสิ่งมีชีวิตทั้งสองและสิ่งมีชีวิตอื่น ๆ ปรากฏการณ์นี้เรียกว่า symbiosis
ต้องขอบคุณ symbiosis กับแบคทีเรียที่เป็นปมทำให้พืชตระกูลถั่วทำให้ดินมีไนโตรเจนเพิ่มขึ้นซึ่งช่วยเพิ่มผลผลิต
การกระจายตัวในธรรมชาติ
จุลินทรีย์มีอยู่ทั่วไปทุกหนทุกแห่ง ข้อยกเว้นเพียงอย่างเดียวคือหลุมอุกกาบาตของภูเขาไฟที่ยังคุกรุ่นและพื้นที่เล็กๆ ที่ศูนย์กลางของระเบิดปรมาณู ทั้งอุณหภูมิต่ำของทวีปแอนตาร์กติกาหรือกระแสน้ำเดือดของกีย์เซอร์หรือสารละลายเกลืออิ่มตัวในสระน้ำเกลือหรือความร้อนแรงของยอดเขาหรือการฉายรังสีอย่างรุนแรงของเครื่องปฏิกรณ์นิวเคลียร์จะรบกวนการดำรงอยู่และการพัฒนาของจุลินทรีย์ สิ่งมีชีวิตทุกชนิดมีปฏิสัมพันธ์กับจุลินทรีย์อยู่ตลอดเวลา ซึ่งไม่เพียงแต่เป็นแหล่งกักเก็บเท่านั้น แต่ยังรวมถึงผู้จัดจำหน่ายด้วย จุลินทรีย์เป็นสิ่งมีชีวิตพื้นเมืองในโลกของเรา โดยกระตือรือร้นในการสำรวจพื้นผิวตามธรรมชาติที่น่าทึ่งที่สุด
จุลินทรีย์ในดิน
จำนวนแบคทีเรียในดินมีขนาดใหญ่มาก - หลายร้อยล้านและพันล้านตัวต่อกรัม มีอยู่ในดินมากกว่าในน้ำและอากาศ จำนวนแบคทีเรียทั้งหมดในดินเปลี่ยนแปลงไป จำนวนแบคทีเรียขึ้นอยู่กับชนิดของดิน สภาพของพวกมัน และความลึกของชั้นดิน
บนพื้นผิวของอนุภาคดิน จุลินทรีย์จะอยู่ในไมโครโคโลนีขนาดเล็ก (เซลล์ละ 20-100 เซลล์) พวกมันมักจะพัฒนาตามความหนาของก้อนอินทรียวัตถุ บนรากพืชที่มีชีวิตและกำลังจะตาย ในเส้นเลือดฝอยบาง ๆ และก้อนภายใน
จุลินทรีย์ในดินมีความหลากหลายมาก มีกลุ่มแบคทีเรียทางสรีรวิทยาที่แตกต่างกัน: แบคทีเรียที่เน่าเปื่อย, แบคทีเรียไนตริไฟดิ้ง, แบคทีเรียตรึงไนโตรเจน, แบคทีเรียซัลเฟอร์ ฯลฯ ในหมู่พวกเขามีแอโรบและแอนแอโรบีสสปอร์และรูปแบบที่ไม่ใช่สปอร์ จุลินทรีย์เป็นปัจจัยหนึ่งในการสร้างดิน
พื้นที่พัฒนาจุลินทรีย์ในดินเป็นบริเวณที่อยู่ติดกับรากของพืชที่มีชีวิต มันถูกเรียกว่าไรโซสเฟียร์และจำนวนจุลินทรีย์ทั้งหมดที่มีอยู่ในนั้นเรียกว่าจุลินทรีย์ไรโซสเฟียร์
จุลินทรีย์ของอ่างเก็บน้ำ
น้ำเป็นสภาพแวดล้อมทางธรรมชาติที่จุลินทรีย์พัฒนาขึ้นเป็นจำนวนมาก ส่วนใหญ่จะลงไปในน้ำจากดิน ปัจจัยที่กำหนดจำนวนแบคทีเรียในน้ำและการมีอยู่ของสารอาหารในน้ำ น้ำที่สะอาดที่สุดมาจากบ่อบาดาลและน้ำพุ อ่างเก็บน้ำและแม่น้ำเปิดอุดมไปด้วยแบคทีเรียมาก แบคทีเรียจำนวนมากที่สุดพบได้ในชั้นผิวน้ำใกล้กับชายฝั่งมากขึ้น เมื่อคุณเคลื่อนออกจากชายฝั่งและเพิ่มความลึก จำนวนแบคทีเรียจะลดลง
น้ำสะอาดมีแบคทีเรีย 100-200 ตัวต่อมิลลิลิตร และน้ำเสียมีแบคทีเรีย 100-300,000 ตัวขึ้นไป มีแบคทีเรียจำนวนมากในตะกอนด้านล่าง โดยเฉพาะในชั้นผิวซึ่งแบคทีเรียจะก่อตัวเป็นแผ่นฟิล์ม ฟิล์มนี้มีแบคทีเรียกำมะถันและเหล็กจำนวนมาก ซึ่งจะออกซิไดซ์ไฮโดรเจนซัลไฟด์เป็นกรดซัลฟิวริก และด้วยเหตุนี้จึงป้องกันไม่ให้ปลาตาย มีรูปแบบที่มีสปอร์มากกว่าในตะกอน ในขณะที่รูปแบบที่ไม่มีสปอร์จะมีอิทธิพลเหนือกว่าในน้ำ
ในแง่ขององค์ประกอบของสายพันธุ์ จุลินทรีย์ของน้ำจะคล้ายกับจุลินทรีย์ในดิน แต่ก็มีรูปแบบเฉพาะเช่นกัน โดยการทำลายของเสียต่างๆ ที่ลงไปในน้ำ จุลินทรีย์จะค่อยๆ ดำเนินการที่เรียกว่าการทำให้น้ำบริสุทธิ์ทางชีวภาพ
จุลินทรีย์ในอากาศ
จุลินทรีย์ในอากาศมีจำนวนน้อยกว่าจุลินทรีย์ในดินและน้ำ แบคทีเรียลอยขึ้นไปในอากาศพร้อมกับฝุ่น สามารถคงอยู่ที่นั่นได้ระยะหนึ่ง จากนั้นจึงตกลงบนพื้นผิวโลกและตายเนื่องจากขาดสารอาหารหรือภายใต้อิทธิพลของรังสีอัลตราไวโอเลต จำนวนจุลินทรีย์ในอากาศขึ้นอยู่กับพื้นที่ทางภูมิศาสตร์ ภูมิประเทศ ช่วงเวลาของปี มลพิษจากฝุ่น ฯลฯ ฝุ่นแต่ละจุดเป็นพาหะของจุลินทรีย์ แบคทีเรียส่วนใหญ่อยู่ในอากาศเหนือสถานประกอบการอุตสาหกรรม อากาศในชนบทก็สะอาดขึ้น อากาศที่สะอาดที่สุดอยู่เหนือป่าไม้ ภูเขา และพื้นที่ที่เต็มไปด้วยหิมะ อากาศชั้นบนมีจุลินทรีย์น้อยลง จุลินทรีย์ในอากาศประกอบด้วยแบคทีเรียที่มีเม็ดสีและมีสปอร์จำนวนมาก ซึ่งมีความทนทานต่อรังสีอัลตราไวโอเลตได้ดีกว่าชนิดอื่นๆ
จุลินทรีย์ของร่างกายมนุษย์
ร่างกายมนุษย์แม้จะมีสุขภาพดีอย่างสมบูรณ์ แต่ก็เป็นพาหะของจุลินทรีย์อยู่เสมอ เมื่อร่างกายมนุษย์สัมผัสกับอากาศและดิน จุลินทรีย์ต่างๆ รวมถึงเชื้อโรค (บาดทะยัก แบคทีเรียเนื้อตายเน่าก๊าซ ฯลฯ) จะเกาะอยู่บนเสื้อผ้าและผิวหนัง ส่วนที่สัมผัสบ่อยที่สุดของร่างกายมนุษย์มีการปนเปื้อน พบเชื้อ E. coli และ staphylococci บนมือ ในช่องปากมีจุลินทรีย์มากกว่า 100 ชนิด ปากซึ่งมีอุณหภูมิ ความชื้น และสารอาหารตกค้าง จึงเป็นสภาพแวดล้อมที่ดีเยี่ยมสำหรับการพัฒนาของจุลินทรีย์
กระเพาะอาหารมีปฏิกิริยาเป็นกรด จุลินทรีย์ส่วนใหญ่ในกระเพาะอาหารจึงตาย เริ่มต้นจากลำไส้เล็กปฏิกิริยาจะกลายเป็นด่างเช่น เป็นผลดีต่อจุลินทรีย์ จุลินทรีย์ในลำไส้ใหญ่มีความหลากหลายมาก ผู้ใหญ่แต่ละคนจะขับถ่ายแบคทีเรียประมาณ 18 พันล้านครั้งต่อวัน เช่น บุคคลมากกว่าคนบนโลก
อวัยวะภายในที่ไม่เชื่อมต่อกับสภาพแวดล้อมภายนอก (สมอง หัวใจ ตับ กระเพาะปัสสาวะ ฯลฯ) มักจะปราศจากจุลินทรีย์ จุลินทรีย์จะเข้าสู่อวัยวะเหล่านี้เฉพาะในช่วงที่เจ็บป่วยเท่านั้น
แบคทีเรียในวัฏจักรของสาร
จุลินทรีย์โดยทั่วไปและโดยเฉพาะอย่างยิ่งแบคทีเรียมีบทบาทสำคัญในวัฏจักรที่มีความสำคัญทางชีวภาพของสารบนโลก โดยทำให้เกิดการเปลี่ยนแปลงทางเคมีที่ไม่สามารถเข้าถึงได้โดยพืชหรือสัตว์โดยสิ้นเชิง ระยะต่างๆ ของวัฏจักรขององค์ประกอบดำเนินไปโดยสิ่งมีชีวิตประเภทต่างๆ การดำรงอยู่ของสิ่งมีชีวิตแต่ละกลุ่มขึ้นอยู่กับการเปลี่ยนแปลงทางเคมีขององค์ประกอบที่ดำเนินการโดยกลุ่มอื่น
วัฏจักรไนโตรเจน
การเปลี่ยนแปลงแบบวงจรของสารประกอบไนโตรเจนมีบทบาทสำคัญในการจัดหาไนโตรเจนในรูปแบบที่จำเป็นให้กับสิ่งมีชีวิตในชีวมณฑลที่มีความต้องการทางโภชนาการที่แตกต่างกัน มากกว่า 90% ของการตรึงไนโตรเจนทั้งหมดเกิดจากกิจกรรมการเผาผลาญของแบคทีเรียบางชนิด
วัฏจักรคาร์บอน
การเปลี่ยนแปลงทางชีวภาพของคาร์บอนอินทรีย์เป็นคาร์บอนไดออกไซด์ ควบคู่ไปกับการลดลงของโมเลกุลออกซิเจน จำเป็นต้องมีกิจกรรมการเผาผลาญร่วมกันของจุลินทรีย์ต่างๆ แบคทีเรียแอโรบิกจำนวนมากทำปฏิกิริยาออกซิเดชันของสารอินทรีย์โดยสมบูรณ์ ภายใต้สภาวะที่ใช้ออกซิเจน สารประกอบอินทรีย์จะถูกสลายขั้นต้นโดยการหมัก และผลิตภัณฑ์อินทรีย์ขั้นสุดท้ายของการหมักจะถูกออกซิไดซ์เพิ่มเติมโดยการหายใจแบบไม่ใช้ออกซิเจน หากมีตัวรับไฮโดรเจนอนินทรีย์ (ไนเตรต ซัลเฟต หรือ CO 2 ) อยู่
วัฏจักรซัลเฟอร์
ซัลเฟอร์มีอยู่ในสิ่งมีชีวิตส่วนใหญ่อยู่ในรูปของซัลเฟตที่ละลายน้ำได้หรือสารประกอบกำมะถันอินทรีย์ที่ลดลง
วัฏจักรเหล็ก
แหล่งน้ำจืดบางแห่งมีเกลือของธาตุเหล็กที่มีความเข้มข้นสูง ในสถานที่ดังกล่าวจุลินทรีย์ในแบคทีเรียโดยเฉพาะจะพัฒนา - แบคทีเรียเหล็กซึ่งออกซิไดซ์ธาตุเหล็กที่ลดลง พวกเขามีส่วนร่วมในการก่อตัวของแร่เหล็กในบึงและแหล่งน้ำที่อุดมไปด้วยเกลือของเหล็ก
แบคทีเรียเป็นสิ่งมีชีวิตที่เก่าแก่ที่สุด ปรากฏตัวเมื่อประมาณ 3.5 พันล้านปีก่อนใน Archean เป็นเวลาประมาณ 2.5 พันล้านปีที่พวกเขาครองโลก ก่อตัวเป็นชีวมณฑล และมีส่วนร่วมในการก่อตัวของชั้นบรรยากาศออกซิเจน
แบคทีเรียเป็นหนึ่งในสิ่งมีชีวิตที่มีโครงสร้างเรียบง่ายที่สุด (ยกเว้นไวรัส) เชื่อกันว่าเป็นสิ่งมีชีวิตชนิดแรกที่ปรากฏบนโลก
พันธุศาสตร์ของแบคทีเรีย
วัตถุประสงค์ของการบรรยาย: เพื่อให้นักเรียนคุ้นเคยกับพื้นฐานของพันธุศาสตร์แบคทีเรียและวิธีการพื้นฐานของการวินิจฉัยยีน
โครงร่างการบรรยาย
1. คุณสมบัติขององค์กรของอุปกรณ์นิวเคลียร์ของแบคทีเรีย
2. องค์ประกอบของจีโนมของแบคทีเรีย
3. ความแปรปรวนของแบคทีเรีย
4. การรวมตัวกันใหม่ของแบคทีเรียและคุณสมบัติของพวกมัน
5. การวินิจฉัยยีน
6. การจำแนกประเภทของแบคทีเรีย
1. คุณสมบัติขององค์กรทางสัณฐานวิทยาของอุปกรณ์นิวเคลียร์ของแบคทีเรีย:
- ไม่มีเยื่อหุ้มนิวเคลียสหรือนิวเคลียส เรียกว่านิวคลอยด์
- DNA เป็นผู้ให้บริการข้อมูลทางพันธุกรรม หากในยูคาริโอต DNA มีลักษณะเป็นเส้นตรง ดังนั้นในแบคทีเรียส่วนใหญ่จะมีลักษณะเป็นวงกลม และมีสายหนึ่งติดอยู่ที่เยื่อหุ้มไซโตพลาสซึม หากคุณคลาย DNA ความยาวของมันจะมากกว่าความยาวของเซลล์หลายร้อยเท่า DNA ของแบคทีเรียนั้นถูกซุปเปอร์คอยล์
- เซลล์แบคทีเรียประกอบด้วยโครโมโซมหนึ่งอัน ได้แก่ แบคทีเรียเป็นสิ่งมีชีวิตเดี่ยว
2. คุณสมบัติทางชีวเคมี.
- DNA ของแบคทีเรียมีองค์ประกอบเหมือนกับ DNA ยูคาริโอต
- ในแบคทีเรีย DNA อาจมีเบสรอง ซึ่งช่วยปกป้อง DNA จากการทำงานของเอนโดนิวคลีเอสของมันเอง
- ในจีโนมของแบคทีเรียที่ทำให้เกิดโรคมีส่วน DNA ที่แตกต่างจากจีโนมหลักในองค์ประกอบคู่เบสนิวคลีโอไทด์ G-C พื้นที่เหล่านี้มีหน้าที่รับผิดชอบในการสังเคราะห์ปัจจัยการทำให้เกิดโรค—หมู่เกาะที่ทำให้เกิดโรค
- DNA ของแบคทีเรียไม่มีฮิสโตน และโพลีเอมีนมีบทบาทในบทบาทของพวกมัน จีโนมของแบคทีเรียแสดงโดยโครงสร้างที่มีความสามารถ
การจำลองแบบออฟไลน์ มีโครงสร้างดังกล่าวอยู่สองโครงสร้าง: โครโมโซมซึ่งมีการเข้ารหัสข้อมูลสำคัญทั้งหมด (โครโมโซมของแบคทีเรียประกอบด้วยยีนที่แตกต่างกันมากถึง 3,000 ยีน) และพลาสมิด
พลาสมิดเป็น DNA ที่มีลักษณะเป็นวงกลม พลาสมิดในเซลล์สามารถอยู่ในสถานะทางเลือกใดสถานะหนึ่งจากสองสถานะ: อิสระหรือรวมเข้ากับโครโมโซม
พลาสมิดเข้ารหัสข้อมูลทางพันธุกรรมเพิ่มเติมที่ไม่สำคัญสำหรับเซลล์ แต่การมีอยู่ของข้อมูลนี้ทำให้มีข้อได้เปรียบในการคัดเลือกบางประการ พลาสมิดได้แก่:
ยีนโครงสร้าง - ยีนที่รับผิดชอบในการจำลองพลาสมิดของพวกมันเอง
พลาสมิดบางชนิดมียีนที่ช่วยให้มั่นใจในการถ่ายทอดของยีนพลาสมิด - ทรา
ตามคุณลักษณะที่เข้ารหัส พวกเขาแยกแยะได้:
- R พลาสมิดเข้ารหัสการดื้อยาของแบคทีเรีย
- พลาสมิด F (เพศ) - กำหนดความสามารถของเซลล์ในการเป็นผู้บริจาคข้อมูลทางพันธุกรรม
- Col plasmid - เข้ารหัสการสังเคราะห์แบคทีเรีย
- พลาสมิดที่รับผิดชอบในการสังเคราะห์ปัจจัยความรุนแรง(Ent-, Hly-)
และพลาสมิดอื่นๆ
จีโนมของแบคทีเรียประกอบด้วยองค์ประกอบทางพันธุกรรมที่เคลื่อนที่ได้: องค์ประกอบ IS (ลำดับการแทรก) ทรานสโพซันและจำนวนเต็ม พบได้ทั้งเป็นส่วนหนึ่งของโครโมโซมของแบคทีเรียและเป็นส่วนหนึ่งของพลาสมิด การจำลองแบบเป็นส่วนสำคัญของการจำลองโครโมโซมและพลาสมิด
องค์ประกอบของ IS เป็นลำดับนิวคลีโอไทด์สั้น (2000) คุณลักษณะที่โดดเด่นขององค์ประกอบ IS คือการมีอยู่ของการซ้ำแบบกลับด้านที่ส่วนท้าย ซึ่งรับรู้โดยทรานสโพเฟส พวกมันไม่มียีนที่มีโครงสร้าง มีความเหมือนกันในแบคทีเรียชนิดต่าง ๆ จำพวก และเชื่อด้วยซ้ำว่าพวกมันเหมือนกันในโปรคาริโอตและยูคาริโอต องค์ประกอบ IS สามารถเคลื่อนที่ได้ทั้งไปตามโครโมโซมและระหว่างโครโมโซม ประกอบด้วย 2 ยีน: ยีนที่ 1 เข้ารหัสการสังเคราะห์ทรานสโพเฟส เอนไซม์นี้ช่วยให้แน่ใจว่ากระบวนการแยกองค์ประกอบ IS ออกจากโครโมโซมและการรวมเข้ากับตำแหน่งของโครโมโซมใหม่ ยีนที่ 2 เข้ารหัสการสังเคราะห์สารกดประสาท ซึ่งควบคุมกระบวนการเคลื่อนไหวทั้งหมด
Transposons คือส่วนของ DNA ที่มีคุณสมบัติเหมือนกับองค์ประกอบ IS แต่มียีนที่มีโครงสร้าง
อินทิกรอนเป็นองค์ประกอบทางพันธุกรรมที่เคลื่อนที่ได้ พวกมันมียีนที่เข้ารหัสการดื้อยาปฏิชีวนะ Integrons เป็นระบบสำหรับจับองค์ประกอบ DNA ขนาดเล็กที่เรียกว่า gene cassette ผ่านการรวมตัวกันใหม่เฉพาะไซต์และการแสดงออกของพวกมัน
ความหมายขององค์ประกอบมือถือ
การเคลื่อนที่ไปตาม DNA ของเซลล์หรือระหว่าง DNA ของเซลล์ต่าง ๆ ทำให้เกิด:
- การปิดใช้งานยีนของส่วนต่าง ๆ ของ DNA ที่มีการเคลื่อนย้ายจะถูกรวมเข้าด้วยกัน
- ความเสียหายต่อสารพันธุกรรม
- การรวมตัวของพลาสมิดเข้ากับโครโมโซม
- การกระจายตัวของยีนในกลุ่มแบคทีเรีย
แบคทีเรียก็เหมือนกับสิ่งมีชีวิตอื่นๆ ที่มีความแปรปรวน การแปรผันของยูคาริโอตเกิดขึ้นในแนวตั้งในแบคทีเรียทั้งในแนวตั้งและแนวนอน
ความแปรปรวนมีสองประเภท: - ฟีโนไทป์ - จีโนไทป์
ความแปรปรวนของฟีโนไทป์แสดงออกมาในรูปแบบของการปรับเปลี่ยน - นี่คือการเปลี่ยนแปลงคุณสมบัติของเซลล์ภายใต้อิทธิพลของอิทธิพลภายนอก
การปรับเปลี่ยนอาจเป็นระยะยาวหรือระยะสั้นก็ได้ การเปลี่ยนแปลงการปรับเปลี่ยนส่งผลต่อเซลล์ส่วนใหญ่ในประชากร
Genotypic คือการกลายพันธุ์หรือการรวมตัวกันใหม่ การกลายพันธุ์อาจเกิดขึ้นเองหรือเกิดขึ้นได้
การรวมตัวกันใหม่คือการทำงานร่วมกันระหว่างจีโนมสองตัวที่มีจีโนไทป์ต่างกัน ซึ่งนำไปสู่การก่อตัวของจีโนมที่รวมยีนของผู้บริจาคและผู้รับเข้าด้วยกัน ในระหว่างกระบวนการรวมตัวกันใหม่ แบคทีเรียจะถูกแบ่งตามอัตภาพออกเป็นเซลล์ผู้บริจาคซึ่งถ่ายโอนสารพันธุกรรมและเซลล์ผู้รับซึ่งรับมัน การรวมตัวกันใหม่ของแบคทีเรียถือว่าคล้ายคลึงกับการสืบพันธุ์แบบอาศัยเพศ
คุณสมบัติของการรวมตัวกันใหม่ในแบคทีเรีย:
- ไม่มีไมโอซิส ไม่ใช่ไซโกตที่เกิดขึ้น แต่เป็นเมราซิโกต
- ส่งตรงจากผู้บริจาคไปยังผู้รับเสมอ
- ปริมาณของสารพันธุกรรมในรีคอมบิแนนท์จะมีมากกว่าหนึ่งเสมอ รีคอมบิแนนท์ประกอบด้วยข้อมูลทางพันธุกรรมทั้งหมดของผู้รับและส่วนหนึ่ง
ข้อมูลทางพันธุกรรมของผู้บริจาค
ในยูคาริโอตมีกลไกการรวมตัวกันใหม่เพียงกลไกเดียวเท่านั้น - ไมโอซิส; ในแบคทีเรียมีการรวมตัวกันใหม่สามประเภท:
1) การเปลี่ยนแปลงคือการแลกเปลี่ยนข้อมูลทางพันธุกรรมโดยใช้ DNA บริสุทธิ์
2) การถ่ายโอนเป็นวิธีการถ่ายโอนข้อมูลทางพันธุกรรมโดยใช้ฟาจ
3) การผันคำกริยาเป็นวิธีการถ่ายโอนข้อมูลทางพันธุกรรมเมื่อมีการสร้างสะพานไซโตพลาสซึมระหว่างแบคทีเรียสองตัว ในระหว่างการผันคำกริยา
จีโนมเกือบทั้งหมดสามารถผ่านเข้าไปในเซลล์ผู้รับได้
วิธีการทางพันธุกรรมใช้เพื่อวัตถุประสงค์ในทางปฏิบัติทั้งในการตรวจหาจุลินทรีย์ในวัสดุภายใต้การศึกษาโดยไม่ต้องแยกวัฒนธรรมบริสุทธิ์ และเพื่อกำหนดตำแหน่งทางอนุกรมวิธานของจุลินทรีย์และดำเนินการระบุตัวตนภายในความจำเพาะ
การจัดลำดับจีโนม– การกำหนดลำดับของคู่นิวคลีโอไทด์ DNA
การวิเคราะห์ข้อจำกัด- วิธีการนี้ขึ้นอยู่กับการใช้เอนไซม์จำกัด - เหล่านี้คือเอ็นโดนิวคลีเอสที่แยกโมเลกุล DNA ในบางสถานที่เท่านั้น หาก DNA ที่แยกได้จากวัสดุเฉพาะได้รับการบำบัดด้วยเอนไซม์จำกัดเฉพาะ สิ่งนี้จะนำไปสู่การก่อตัวของชิ้นส่วน DNA ในขนาดคงที่ตามจำนวนที่กำหนดอย่างเคร่งครัด
การพิมพ์ริบบิ้น– ช่วยให้คุณกำหนดชนิดของแบคทีเรียได้ ลำดับของเบสนิวคลีโอไทด์ในโอเปอรอนที่เข้ารหัส rRNA นั้นมีลักษณะพิเศษคือการมีอยู่ของทั้งสองบริเวณที่ได้รับการอนุรักษ์ ซึ่งมีโครงสร้างคล้ายกันในแบคทีเรียที่แตกต่างกัน และลำดับที่แปรผันได้ ซึ่งเป็นสกุลและจำเพาะต่อสปีชีส์ และเป็นเครื่องหมายสำหรับการจำแนกทางพันธุกรรม
การผสมพันธุ์ระดับโมเลกุล– ใช้ในการจัดระบบยีน วิธีนี้ช่วยให้เราสามารถกำหนดระดับความคล้ายคลึงกันระหว่าง DNA ต่างๆ ได้
PCR ใช้เพื่อตรวจจับยีนหรือลำดับนิวคลีโอไทด์ที่เกี่ยวข้องซึ่งเข้ารหัสสปีชีส์หรือลักษณะอื่น
วิธี PCR ขึ้นอยู่กับหลักการเสริมและช่วยให้คุณสามารถเพิ่ม (ขยาย) จำนวนตัวอย่าง DNA ที่กำลังทดสอบได้ วิธีนี้มีความไวสูงมาก และในทางทฤษฎีทำให้สามารถตรวจพบโมเลกุล DNA เดี่ยวในวัสดุที่กำลังศึกษาได้
ประเภทของ PCR:
- PCR แบบเรียลไทม์; ทำให้สามารถระบุจำนวนชิ้นส่วน DNA ที่มีอยู่ในวัสดุได้เช่น ดำเนินการวิเคราะห์เชิงปริมาณ
- multiplex PCR - ข้อดีคือสามารถใส่ไพรเมอร์ 2-4 คู่ขึ้นไปลงในส่วนผสมของปฏิกิริยาได้ เป็นลักษณะของเชื้อโรคต่างๆ
- Reverse Transcription PCR – ช่วยให้สามารถคัดลอก RNA ของเชื้อโรคได้
ชิป DNA เป็นเทคโนโลยีล่าสุดสำหรับวิธีไฮบริดในการวิเคราะห์ทางอณูพันธุศาสตร์ พวกมันเป็นพาหะของโอลิโกนิวคลีโอไทด์ที่รู้จัก (น้อยกว่า 20 เบสในแต่ละเบส) ซึ่งประกอบกับส่วนของจีโนม (หรือจีโนม) ที่กำลังศึกษาและครอบครองพื้นที่เฉพาะ (เซลล์) หากมีชิ้นส่วนของ DNA ที่ต้องการในตัวอย่างทดสอบ พวกมันจะผสมกัน (เชื่อมต่อตามหลักการเสริมกัน) กับลำดับนิวคลีโอไทด์ที่อยู่บนชิป
การจำแนกประเภทของแบคทีเรีย
หน่วยอนุกรมวิธานพื้นฐานของแบคทีเรียคือสายพันธุ์ ในการระบุชนิดพันธุ์ในแบคทีเรีย จะใช้ระบบการตั้งชื่อแบบคู่ (ไบนารี่)
สายพันธุ์ในแบคทีเรียคือกลุ่มของแบคทีเรียที่เกี่ยวข้องซึ่งมีคุณสมบัติทางชีวภาพคล้ายคลึงกันและมีต้นกำเนิดร่วมกัน ปัจจุบันมี 3 วิธีในการจำแนกแบคทีเรีย:
1. การจำแนกประเภทประจำ
เป็นพื้นฐานของคำแนะนำเกี่ยวกับแบคทีเรียที่แก้ไขโดย Bergey
2. อนุกรมวิธานเชิงตัวเลข
3. ระบบยีน
สรุป: นักเรียนมีความคุ้นเคยกับพื้นฐานของพันธุศาสตร์ของแบคทีเรียและวิธีการพื้นฐานของการวินิจฉัยยีน
ไวรัสที่ประกอบด้วย DNA มีเอนไซม์การจำลองของตัวเอง (ใน capsid) หรือจีโนมของพวกมันเข้ารหัสข้อมูลเกี่ยวกับการสังเคราะห์เอนไซม์ของไวรัสที่รับรองการจำลองกรดนิวคลีอิกของไวรัส ปริมาณของเอนไซม์เหล่านี้จะแตกต่างกันไปเมื่อนำไปใช้กับไวรัสต่างๆ ตัวอย่างเช่น จีโนมของไวรัสแบคทีเรีย T4 เข้ารหัสข้อมูลเกี่ยวกับการสังเคราะห์เอนไซม์ของไวรัสประมาณ 30 ชนิด นอกจากนี้จีโนมของไวรัสขนาดใหญ่ยังเข้ารหัสนิวคลีเอสที่ทำลาย DNA ของเซลล์เจ้าบ้าน เช่นเดียวกับโปรตีน ซึ่งผลกระทบต่อ RNA polymerase ของเซลล์นั้นมาพร้อมกับความจริงที่ว่า "RNA polymerase ที่ได้รับการบำบัดด้วยวิธีนี้จะถ่ายทอดยีนของไวรัสที่แตกต่างกัน ในระยะต่างๆ ของการติดเชื้อไวรัส ในทางตรงกันข้าม ไวรัส DNA ขนาดเล็กจะขึ้นอยู่กับเอนไซม์ของเซลล์เจ้าบ้านมากกว่า ตัวอย่างเช่น การสังเคราะห์ DNA ของอะดีโนไวรัสนั้นมั่นใจได้ด้วยเอนไซม์ของเซลล์[...]
DNA ของแบคทีเรียเป็นสารประกอบโพลีเมอร์สูงที่ประกอบด้วยนิวคลีโอไทด์จำนวนมาก - โพลีนิวคลีโอไทด์ที่มีน้ำหนักโมเลกุลประมาณ 4 ล้าน โมเลกุล DNA เป็นสายโซ่ของนิวคลีโอไทด์ซึ่งการจัดเรียงของพวกมันมีลำดับที่แน่นอน ลำดับของเบสไนโตรเจนจะเข้ารหัสข้อมูลทางพันธุกรรมของแต่ละสปีชีส์ การละเมิดลำดับนี้อาจเกิดขึ้นได้เนื่องจากการกลายพันธุ์ตามธรรมชาติหรือภายใต้อิทธิพลของปัจจัยก่อกลายพันธุ์ ในกรณีนี้จุลินทรีย์จะได้หรือสูญเสียทรัพย์สินบางส่วนไป ลักษณะของมันเปลี่ยนไปตามกรรมพันธุ์นั่นคือจุลินทรีย์รูปแบบใหม่ปรากฏขึ้น ในจุลินทรีย์ทั้งหมด - โปรคาริโอตและยูคาริโอต - พาหะของข้อมูลทางพันธุกรรมคือกรดนิวคลีอิก - DNA และ RNA มีเพียงไวรัสบางชนิดเท่านั้นที่เป็นข้อยกเว้น เนื่องจากไม่มี DNA และข้อมูลทางพันธุกรรมจะถูกบันทึกหรือสะท้อนให้เห็นเฉพาะใน RNA[...]
ในเซลล์แบคทีเรีย จำนวนฐาน DNA ทั้งหมดประกอบด้วยกัวนีนและไซโตซีน 32-65 โมล%[...]
นิวเคลียสของเซลล์แบคทีเรีย ประมาณ 1-2% ของมวลแห้งของจุลินทรีย์คือ DNA ซึ่งมีข้อมูลทางพันธุกรรมของสิ่งมีชีวิต จุลินทรีย์ส่วนใหญ่มีพื้นที่ (หรือหลายพื้นที่) ซึ่ง DNA จำนวนมากมีความเข้มข้น มีโครงสร้างเฉพาะ (หรือออร์แกเนลล์) และเรียกว่านิวเคลียส นิวเคลียส (หรือสารนิวเคลียร์) มีความเกี่ยวข้องกับเยื่อหุ้มเซลล์ไซโตพลาสซึม โดยไม่คำนึงว่าจะถูกล้อมรอบด้วยเยื่อหุ้มพื้นฐาน (เช่นในอะมีบา) หรือไม่มีเยื่อหุ้มเหล่านั้น (เช่นในแบคทีเรียและสาหร่ายสีเขียวแกมน้ำเงิน) สารนิวเคลียร์จะถูกกระตุ้นในช่วงการสืบพันธุ์และเมื่อเริ่มมีการเปลี่ยนแปลงที่เกี่ยวข้องกับอายุที่เกี่ยวข้องกับการแก่ชราของเซลล์[...]
ส่วน DNA (ยีน) ที่มีไว้สำหรับการโคลนระดับโมเลกุลจะต้องมีความสามารถในการทำซ้ำเมื่อถูกถ่ายโอนไปยังเซลล์แบคทีเรีย กล่าวคือ เป็นแบบจำลอง อย่างไรก็ตามเขาไม่มีความสามารถเช่นนั้น ดังนั้นเพื่อให้แน่ใจว่ามีการถ่ายโอนและตรวจจับยีนที่โคลนในเซลล์ พวกมันจึงถูกรวมเข้ากับสิ่งที่เรียกว่าพาหะทางพันธุกรรม อย่างหลังจะต้องมีคุณสมบัติอย่างน้อยสองประการ ขั้นแรก เวกเตอร์จะต้องสามารถทำซ้ำในเซลล์และหลายสำเนาได้ ประการที่สอง พวกเขาจะต้องจัดให้มีความสามารถในการเลือกเซลล์ที่มีเวกเตอร์ กล่าวคือ มีเครื่องหมายที่สามารถใช้เพื่อตอบโต้การเลือกเซลล์ที่มีเวกเตอร์พร้อมกับยีนที่ถูกโคลน (โมเลกุล DNA ลูกผสม) พลาสมิดและฟาจเป็นไปตามข้อกำหนดเหล่านี้ พลาสมิดเป็นพาหะที่ดีเนื่องจากเป็นแบบจำลองและสามารถมียีนที่ต้านทานต่อยาปฏิชีวนะใด ๆ ซึ่งช่วยให้สามารถเลือกแบคทีเรียที่จะต้านทานต่อยาปฏิชีวนะนี้ได้ดังนั้นจึงสามารถตรวจจับโมเลกุล DNA ลูกผสมได้ง่าย [...]
ในแบคทีเรีย DNA จะถูกอัดแน่นน้อยกว่า ไม่เหมือนนิวเคลียสที่แท้จริง นิวคลอยด์ไม่มีเมมเบรน นิวคลีโอลัส หรือชุดโครโมโซม DNA ของแบคทีเรียไม่เกี่ยวข้องกับโปรตีนหลัก - ฮิสโตน - และตั้งอยู่ในนิวเคลียสในรูปแบบของมัดของไฟบริล[...]
การใช้เทคนิครีคอมบิแนนท์ DNA ในการผลิตสารชีวภาพสำหรับการควบคุมมลพิษยังอยู่ในระยะเริ่มต้น แต่เทคนิคหนึ่งที่อาจพิสูจน์ได้ว่ามีประโยชน์ในอนาคตอันใกล้ก็คือการตรวจสอบทางพันธุกรรม การคัดเลือกสิ่งมีชีวิตที่สามารถเปลี่ยนรูปสารประกอบใหม่ได้มักขึ้นอยู่กับความสามารถในการใช้สารดังกล่าวเป็นสารตั้งต้นในการเจริญเติบโต หากการเจริญเติบโตอ่อนแอหรือสารตั้งต้นถูกเผาผลาญเท่านั้น วิธีการคัดเลือกจะไม่เหมาะสำหรับการระบุความสามารถในการย่อยสลาย ดังนั้นจึงจะเป็นประโยชน์ในการพัฒนาการตรวจทางพันธุกรรมเพื่อระบุลำดับเฉพาะในพลาสมิดและโครโมโซม ซึ่งเป็นสิ่งจำเป็นในการพิจารณาศักยภาพของแคทาบอลิซึม แม้ว่าศักยภาพนี้จะไม่ได้แสดงออกมาก็ตาม โพรบดังกล่าวได้รับการออกแบบสำหรับพลาสมิดของ TOL วิธีการนี้สามารถระบุโคโลนีของแบคทีเรียหนึ่งกลุ่มที่มีพลาสมิด TOL อยู่ในโคโลนี Escherichia coli 106 โคโลนี เครื่องมืออันทรงพลังดังกล่าวจะมีคุณค่าอย่างยิ่งในการแยกฟังก์ชัน catabolic ที่ซ่อนอยู่[...]
การพัฒนาเทคนิคอันหรูหราสำหรับการ "โคลนนิ่ง" DNA เพื่อสร้างสำเนาชิ้นส่วน DNA เฉพาะเจาะจงจำนวนมาก (รูปที่ 13.4) เพิ่งเปิดโลกทัศน์ใหม่ในการศึกษาโครงสร้าง การจัดองค์กร และหน้าที่ของจีโนม หากคุณแยก DIC แบบเกลียวคู่ออกด้วยเอนไซม์ "จำกัด" ตัวใดตัวหนึ่ง (หนึ่งในนิวเคลียส) ซึ่งจดจำและแยกลำดับนิวคลีโอไทด์สั้น ๆ (4-6 คู่) โดยเฉพาะ จากนั้นชิ้นส่วน DNA ที่สามารถทำซ้ำได้สูงจะปรากฏขึ้น ปลายของสาย DNA ทั้งสองมักจะถูกแทนที่โดยสัมพันธ์กันเนื่องจากความจำเพาะของบริเวณที่ตัดของโมเลกุลที่มีเกลียวคู่ ซึ่งสายดังกล่าวเป็นส่วนเสริมในองค์ประกอบฐาน โดยปกติแล้ว DNA จะถูกแทรกเข้าไปในยีนพลาสมิดที่มีความสำคัญต่อการผสมพันธุ์ เช่น ยีนต้านทานยาปฏิชีวนะ ซึ่งช่วยให้แบคทีเรียที่มีพลาสมิดเติบโตได้เมื่อมียาปฏิชีวนะ[...]
ในแบคทีเรีย การจำลองแบบจะสร้างสำเนาของพลาสมิดจำนวนมาก และด้วยวิธีนี้ ชิ้นส่วน DNA ที่ฝังอยู่จำนวนมากจึงสามารถ "เติบโต" ได้ จากนั้นจึงแยกออกอีกครั้งโดยการย่อยด้วยเอนไซม์จำกัดเดียวกัน และการแยกผลิตภัณฑ์ที่เป็นผลลัพธ์โดยเจลอิเล็กโตรโฟรีซิส การใช้วิธีรวมตัวใหม่ของ DNA นี้ได้ปฏิวัติการศึกษายีน[...]
เพิ่งค้นพบว่ารังสีความเข้มต่ำที่มีความยาวคลื่น 320-400 นาโนเมตร (บริเวณใกล้กับโซนแสงที่มองเห็นได้) มีผลกระทบต่อการกลายพันธุ์ในไวรัส DNA ของแบคทีเรีย ยังไม่มีการค้นพบผลกระทบที่เป็นไปได้ของการแผ่รังสีในช่วงความยาวคลื่นนี้ต่อไวรัสพืช[...]
เส้นโค้งของการพึ่งพาการเชื่อมโยงใหม่ของ COT ที่ได้รับสำหรับ DNA ของแบคทีเรียนั้นไม่มีข้อบกพร่องและ DIC ของยูคาริโอตจะเชื่อมโยงใหม่ตามประเภทที่แตกต่างกัน (รูปที่ 13.2) ที่ความเข้มข้นของ DNA ต่ำและเวลาฟักตัวสั้น สัดส่วนที่มีนัยสำคัญของ DNA สายเดี่ยวจะถูกทำให้สุกอีกครั้ง และด้วย COT ที่เพิ่มขึ้น ก็จะเกิดโมเลกุลสายคู่เพิ่มเติมขึ้น เพื่อให้ได้เส้นโค้งสองเฟส . การคืนสภาพอย่างรวดเร็วที่ค่า COT ต่ำ แสดงให้เห็นว่าบางลำดับในยูคาริโอตถูกทำซ้ำหลายครั้ง เช่น มากถึง 10,000 ครั้งขึ้นไป [...]
การไม่มี CXC ยังสามารถเลียนแบบได้ในกรณีที่ DNA ของฟาจทดสอบไม่มีตำแหน่งที่ได้รับการยอมรับจากเอนไซม์ควบคุมที่มีอยู่ในสายพันธุ์ที่กำลังศึกษาอยู่ ปรากฏการณ์นี้แสดงถึงการเปลี่ยนแปลงทางวิวัฒนาการในไวรัสแบคทีเรียที่ออกแบบมาเพื่อช่วยให้พวกเขาเอาชนะอุปสรรค CXC ผลกระทบของแรงกดดันในการคัดเลือกในกรณีนี้จะแสดงออกมาในจำนวนที่ลดลงอย่างมีนัยสำคัญหรือแม้กระทั่งการกำจัดโดยสิ้นเชิงในฟาจ DNA ของลำดับนิวคลีโอไทด์ที่เป็นสารตั้งต้นของลักษณะเฉพาะของเอนไซม์จำกัดของเซลล์เจ้าบ้านของไวรัสแบคทีเรีย[.. .]
Lindegren อธิบายขั้นตอนที่เป็นไปได้ของการก่อตัวของแบคทีเรียแบคเทอริโอฟาจจาก DNA ของผู้ทำนาย โดยบอกว่าการพยากรณ์เกิดขึ้นโดยเป็นส่วนหนึ่งของ DNA ของแบคทีเรียแปลกปลอมที่เข้าไปในเซลล์โดยไม่ได้ตั้งใจ ซึ่งในระยะแรกจะแบ่งตัวพร้อมกันกับ DNA ของแบคทีเรีย ขั้นตอนสำคัญต่อไปในการพัฒนาของไวรัสคือการเปลี่ยนแปลงในการพยากรณ์ซึ่งเป็นผลมาจากการสืบพันธุ์โดยไม่ขึ้นกับ DNA ของเซลล์เจ้าบ้าน ผลก็คือคำทำนายจะถูกนำมาใช้ นิวคลีโอไทด์ที่มีอยู่ทั้งหมด จึงขัดขวางการเจริญเติบโตของเซลล์เจ้าบ้าน ในที่สุด ในระยะต่อมา เปลือกโปรตีนป้องกันอาจก่อตัวขึ้นและโปรตีนอื่นๆ ก็จะเกิดขึ้น ซึ่งรับประกันความอยู่รอดของ DNA ภายนอกร่างกายของโฮสต์และการติดเชื้อเซลล์ใหม่อย่างมีประสิทธิภาพ ชิ้นส่วนที่แยกออกจากกันของ DNA ของแบคทีเรียเริ่มแรกเห็นได้ชัดว่ามีการเข้ารหัสโปรตีนซึ่งปรับให้เข้ากับการทำงานของแบคทีเรีย การเปลี่ยนแปลงที่สำคัญมากใน DNA เป็นสิ่งจำเป็นสำหรับวัตถุที่ซับซ้อนและมีความเชี่ยวชาญ เช่น T2 phage ของ E. coli ซึ่งมีเบสที่ไม่มีอยู่ใน DIC ของแบคทีเรียด้วยที่จะเกิดขึ้น [...]
ข้อมูลทางพันธุกรรมของแบคทีเรียไม่ได้จำกัดอยู่เพียง DNA ที่อยู่ในนิวเคลียสของเซลล์แบคทีเรีย ดังที่ได้กล่าวไว้แล้วในส่วนก่อนหน้าของหนังสือ องค์ประกอบนอกโครโมโซมซึ่งเรียกรวมกันว่าพลาสมิดก็ทำหน้าที่เป็นพาหะของคุณสมบัติทางพันธุกรรมเช่นกัน นิวคลอยด์ซึ่งเป็นออร์แกเนลล์ของเซลล์แบคทีเรียต่างจาก DNA นิวเคลียร์ที่เทียบเท่ากัน พลาสมิดเป็นองค์ประกอบทางพันธุกรรมที่เป็นอิสระ การสูญเสียพลาสมิดหรือการได้มานั้นไม่ส่งผลกระทบต่อชีววิทยาของเซลล์ (การได้มาของพลาสมิดมีผลเชิงบวกต่อประชากรโดยรวมเท่านั้น ซึ่งเพิ่มความมีชีวิตของสายพันธุ์) พลาสมิดที่ถ่ายทอดได้รวมถึงพลาสมิดที่เริ่มคุณสมบัติของผู้บริจาคในเซลล์เจ้าบ้าน ในเวลาเดียวกันสิ่งหลังได้รับคุณภาพใหม่ - ความสามารถในการเชื่อมต่อกับเซลล์ผู้รับและให้พลาสมิดแก่พวกมัน เซลล์ผู้รับ ซึ่งได้รับพลาสมิดระหว่างการผันคำกริยา ตัวเองจะกลายเป็นผู้บริจาค[...]
การไม่มีการดูดซึมไม่ได้ทำให้ทางเลือกอันหลากหลายระหว่างไวรัสแบคทีเรียและเซลล์จุลินทรีย์หมดไป พวกเขาแสดงให้เห็นเพียงด้านเดียวของปรากฏการณ์นี้ กล่าวคือ การปรากฏของกลไกการปกป้องเซลล์ที่เลียนแบบข้อจำกัดทางฟีโนไทป์ (ตามเกณฑ์ของการขาดการเติบโต) อย่างไรก็ตาม มีปฏิสัมพันธ์ระหว่างเซลล์และแบคทีเรียอีกรูปแบบหนึ่งที่สามารถเลียนแบบการไม่มี CXC ตัวอย่างของกลไกดังกล่าวคือการสังเคราะห์สารยับยั้งและเมทิเลสที่เข้ารหัสโดยยีนฟาจที่ปกป้อง DNA ของไวรัสจากการทำงานของเอนไซม์จำกัดประเภท II[...]
กลไกการออกฤทธิ์ในการฆ่าเชื้อของคลอรีนสัมพันธ์กับความผิดปกติของการเผาผลาญของเซลล์แบคทีเรียในระหว่างกระบวนการฆ่าเชื้อในน้ำ ในเวลาเดียวกัน มีการเผยให้เห็นถึงผลกระทบต่อการทำงานของเอนไซม์ของแบคทีเรีย โดยเฉพาะอย่างยิ่งต่อดีไฮโดรจีเนสที่กระตุ้นปฏิกิริยารีดอกซ์ในเซลล์แบคทีเรีย A. M. Skidalskaya (1969) ศึกษาผลของคลอรีนต่อกระบวนการดีคาร์บอกซิเลชันของกรดอะมิโนจากแบคทีเรียซึ่งเกิดขึ้นต่อหน้าเอนไซม์ดีคาร์บอกซิเลสที่จำเพาะเจาะจงอย่างเคร่งครัดและยังพิจารณาองค์ประกอบนิวคลีโอไทด์ของ Escherichia coli DNA หลังจากเสร็จสิ้นกระบวนการฆ่าเชื้อในระดับต่างๆ มีฤทธิ์ฆ่าเชื้อแบคทีเรีย[ ..]
แบคทีเรียกลุ่ม T มีรูปร่างเหมือนไม้ตีกลองขนาด 100 x 25 นาโนเมตร จีโนมของพวกมันแสดงโดย DNA พวกมันเป็นฟาจที่มีความรุนแรงเนื่องจากหลังจากที่พวกมันติดเชื้อในเซลล์แบคทีเรีย แบคทีเรียตัวหลังจะถูก lysed และปล่อยอนุภาคฟาจที่สังเคราะห์ขึ้นใหม่จำนวนมาก [...]
พลาสมิดของแบคทีเรียเป็นโครงสร้างทางพันธุกรรมที่อยู่ในไซโตพลาสซึมและเป็นตัวแทนของโมเลกุล DNA ซึ่งมีขนาดตั้งแต่ 2,250 ถึง 400,000 คู่เบสไนโตรเจน พวกมันมีอยู่แยกจากโครโมโซมในปริมาณตั้งแต่หนึ่งถึงหลายสิบสำเนาต่อเซลล์แบคทีเรีย[...]
สายพันธุ์ Pseu.dom.onas vug1 ae ri pka8eoIco1a มีความยาวพลาสมิด 150,000 bp ซึ่งสามารถทำซ้ำได้โดยอัตโนมัติหรือสามารถรวมเข้ากับโครโมโซมของแบคทีเรียได้ การตัดออกที่ไม่แม่นยำในเวลาต่อมาทำให้เกิดตระกูลพลาสมิดที่มีความยาวตั้งแต่ 35 ถึง 270 kb ซึ่งบางส่วนมีโครโมโซม DNA ขนาดใหญ่[...]
ในระหว่างวิวัฒนาการ แบคทีเรียได้พัฒนาความสามารถในการสังเคราะห์สิ่งที่เรียกว่าเอนไซม์ควบคุม (เอนโดนิวคลีเอส) ซึ่งต่อมาได้กลายเป็นส่วนหนึ่งของระบบปรับเปลี่ยนข้อจำกัดของเซลล์ (แบคทีเรีย) ในแบคทีเรีย ระบบจำกัดการปรับเปลี่ยนคือระบบภูมิคุ้มกันภายในเซลล์สำหรับการป้องกัน DNA แปลกปลอม ซึ่งแตกต่างจากสิ่งมีชีวิตที่สูงขึ้นซึ่งการรับรู้และการทำลายไวรัสแบคทีเรียและเชื้อโรคอื่น ๆ เกิดขึ้นนอกเซลล์ในแบคทีเรียการป้องกันจาก DNA แปลกปลอม (DNA ของพืชและสัตว์ในร่างกายที่พวกมันอาศัยอยู่) เกิดขึ้นในเซลล์เช่น เมื่อ DNA แปลกปลอมแทรกซึมเข้าไปในไซโตพลาสซึม ของแบคทีเรีย เพื่อปกป้องตัวเอง แบคทีเรียยังได้พัฒนาความสามารถในการ "แท็ก" DNA ของตัวเองด้วยเบสเมทิลเลชันในลำดับบางอย่าง ด้วยเหตุนี้ DNA แปลกปลอมเนื่องจากการไม่มีกลุ่มเมทิลในลำดับเดียวกัน จึงถูกละลาย (ตัด) เป็นชิ้นส่วนโดยเอนไซม์จำกัดแบคทีเรียหลายชนิด จากนั้นจึงสลายตัวโดยเอ็กโซนิวคลีเอสของแบคทีเรียเป็นนิวคลีโอไทด์ เราสามารถพูดได้ว่าด้วยวิธีนี้แบคทีเรียจึงป้องกันตัวเองจาก DNA ของพืชและสัตว์ ซึ่งพวกมันอาศัยอยู่ในร่างกายชั่วคราว (เป็นเชื้อโรค) หรือถาวร (เป็น saprophytes)[...]
คุณสมบัติทางพันธุกรรมของแบคทีเรียหรือลักษณะเฉพาะของแต่ละบุคคลจะถูกเข้ารหัสในหน่วยพันธุกรรม - ยีนที่อยู่ในโครโมโซมตามแนวเส้นดีเอ็นเอ ด้วยเหตุนี้ ยีนจึงเป็นส่วนหนึ่งของสาย DNA ลักษณะเฉพาะแต่ละอย่างสอดคล้องกับยีนเฉพาะและมักจะเป็นโคดอนชิ้นเล็ก ๆ ของ DNA กล่าวอีกนัยหนึ่ง สาย DNA มีข้อมูลเกี่ยวกับคุณสมบัติทั้งหมดของแบคทีเรียในลำดับเชิงเส้น อย่างไรก็ตาม แบคทีเรียยังมีคุณสมบัติอีกอย่างหนึ่ง นิวเคลียสของยูคาริโอตมักจะมีโครโมโซมหลายโครโมโซม จำนวนโครโมโซมในนิวเคลียสจะคงที่ในแต่ละสปีชีส์ นิวเคลียสของแบคทีเรียประกอบด้วยวงแหวน DNA เพียงวงเดียว กล่าวคือ โครโมโซมเดียว อย่างไรก็ตาม ผลรวมของลักษณะทางพันธุกรรมของเซลล์แบคทีเรียไม่ได้หมดไปจากการจัดเก็บข้อมูลที่มีอยู่ในโครโมโซมเดียวหรือในเกลียว DNA ที่มีเกลียวคู่แบบปิดรูปวงแหวน พลาสมิดประกอบด้วย DNA ซึ่งนำข้อมูลทางพันธุกรรมที่ถ่ายทอดจากเซลล์แม่ไปยังเซลล์ลูก[...]
การกลายพันธุ์คือการเปลี่ยนแปลงในอุปกรณ์ยีนของเซลล์ ซึ่งมาพร้อมกับการเปลี่ยนแปลงในลักษณะที่ควบคุมโดยยีนเหล่านี้ มีความเสียหายทั้งในระดับมหภาคและระดับจุลภาคต่อ DNA ซึ่งนำไปสู่การเปลี่ยนแปลงคุณสมบัติของเซลล์ การเปลี่ยนแปลงระดับมหภาค ได้แก่ การสูญเสียส่วนของ DNA (การแบ่งส่วน) การเคลื่อนที่ของส่วนที่แยกจากกัน (การโยกย้าย) หรือการหมุนของส่วนของโมเลกุล 180° (การผกผัน) มักพบในแบคทีเรียค่อนข้างน้อย Microdamages หรือการกลายพันธุ์แบบจุด กล่าวคือ เป็นเรื่องปกติสำหรับพวกเขามากกว่ามาก การเปลี่ยนแปลงเชิงคุณภาพในยีนแต่ละตัว เช่น การแทนที่ฐานไนโตรเจนคู่หนึ่ง การกลายพันธุ์อาจเป็นแบบตรงและแบบผกผันหรือแบบย้อนกลับก็ได้ การกลายพันธุ์โดยตรงคือการกลายพันธุ์ในสิ่งมีชีวิตประเภทป่า ตัวอย่างเช่น การสูญเสียความสามารถในการสังเคราะห์ปัจจัยการเจริญเติบโตอย่างอิสระ เช่น การเปลี่ยนจากโปรโต- ไปเป็นออโซโทรฟี การกลายพันธุ์แบบย้อนกลับแสดงถึงการกลับมาหรือการพลิกกลับไปสู่ประเภทไวด์ ความสามารถในการเปลี่ยนกลับเป็นลักษณะของการกลายพันธุ์แบบจุด อันเป็นผลมาจากการกลายพันธุ์ลักษณะที่สำคัญเช่นความสามารถในการสังเคราะห์กรดอะมิโนและวิตามินอย่างอิสระ (การกลายพันธุ์ของ auxotrophic) และความสามารถในการสร้างการเปลี่ยนแปลงของเอนไซม์ การกลายพันธุ์เหล่านี้เรียกว่าชีวเคมี การกลายพันธุ์ที่นำไปสู่การเปลี่ยนแปลงความไวต่อยาปฏิชีวนะและสารต้านจุลชีพอื่นๆ ก็เป็นที่ทราบกันดีเช่นกัน ขึ้นอยู่กับต้นกำเนิด การกลายพันธุ์แบ่งออกเป็นแบบที่เกิดขึ้นเองและแบบเหนี่ยวนำ เกิดขึ้นเองตามธรรมชาติโดยไม่มีการแทรกแซงของมนุษย์และเกิดขึ้นแบบสุ่มในธรรมชาติ ความถี่ของการกลายพันธุ์ดังกล่าวต่ำมากและอยู่ในช่วงตั้งแต่ 1 X 10"4 ถึง 1 X 10-10 สิ่งกระตุ้นเกิดขึ้นเมื่อจุลินทรีย์สัมผัสกับปัจจัยก่อกลายพันธุ์ทางกายภาพหรือทางเคมี ปัจจัยทางกายภาพที่มีผลกระทบต่อการกลายพันธุ์ ได้แก่ รังสีอัลตราไวโอเลตและรังสีไอออไนซ์ รวมถึงอุณหภูมิ สารประกอบจำนวนหนึ่งเป็นสารก่อกลายพันธุ์ทางเคมี และหนึ่งในสารประกอบเหล่านั้นที่มีฤทธิ์มากที่สุดคือสิ่งที่เรียกว่าซูเปอร์มิวทาเจน ภายใต้สภาพธรรมชาติและในการทดลอง การเปลี่ยนแปลงองค์ประกอบของประชากรแบคทีเรียสามารถเกิดขึ้นได้อันเป็นผลมาจากการกระทำของปัจจัยสองประการ - การกลายพันธุ์และการเลือกอัตโนมัติ ซึ่งเกิดขึ้นอันเป็นผลมาจากการปรับตัวของสายพันธุ์กลายบางชนิดให้เข้ากับสภาพแวดล้อม กระบวนการนี้สังเกตได้อย่างชัดเจนในสภาพแวดล้อมที่แหล่งอาหารหลักคือสารสังเคราะห์ เช่น สารลดแรงตึงผิวหรือคาโปรแลคตัม[...]
เซลล์ E. coli เซลล์เดียวล้อมรอบด้วยเยื่อหุ้มเซลล์สามชั้นหนาประมาณ 40 นาโนเมตร ซึ่งเป็น "ถุง" หรือ "ซอง" ที่บรรจุเนื้อหาของเซลล์ในรูปของโปรตีนประมาณ 2 x 10 16 กรัม 6 x 10 16 กรัมของ DNA และ 2 x 10 14 กรัมของ RNA (ส่วนใหญ่เป็นไรโบโซม RNA) โปรตีนประมาณ 2,000 ชนิดถูกสังเคราะห์ขึ้นในเซลล์แบคทีเรีย ซึ่งส่วนใหญ่พบได้ในไซโตพลาสซึม ความเข้มข้นของโปรตีนบางชนิดคือ 10"® M ในขณะที่โปรตีนบางชนิดมีค่าประมาณ 2 x 10"4 M (ตั้งแต่ 10 ถึง 200,000 โมเลกุลต่อเซลล์)[...]
ในสิ่งมีชีวิตเซลล์เดียว การสืบพันธุ์แบบอาศัยเพศเกิดขึ้นได้หลายรูปแบบ การผันคำกริยายังพบได้ในซิเลียต ซึ่งในระหว่างกระบวนการนี้ การถ่ายโอนนิวเคลียสเกิดขึ้นจากบุคคลหนึ่งไปยังอีกบุคคลหนึ่ง ตามด้วยการแบ่งนิวเคลียสของนิวเคลียสอย่างหลัง[...]
แบคทีเรีย: โปรคาริโอต (“พรีนิวเคลียร์”) สิ่งมีชีวิตเซลล์เดียว เซลล์ของพวกเขาไม่มีนิวเคลียสแยกออกจากไซโตพลาสซึม อย่างไรก็ตาม โปรแกรมทางพันธุกรรม เช่นเดียวกับสิ่งมีชีวิตทั้งหมด ได้รับการเข้ารหัสเป็นลำดับของนิวคลีโอไทด์ใน DNA และนำข้อมูลเกี่ยวกับโครงสร้างของโปรตีน เซลล์แบคทีเรียไม่มีออร์แกเนลล์ เช่น คลอโรพลาสต์ (เฉพาะสำหรับการสังเคราะห์ด้วยแสง) และไมโตคอนเดรีย (เฉพาะสำหรับการหายใจของเซลล์และการสังเคราะห์ ATP) กระบวนการทางชีวเคมีเหล่านี้เกิดขึ้นในแบคทีเรียในไซโตพลาสซึม[...]
ขนาดเซลล์ที่เล็กมากเป็นลักษณะเฉพาะ แต่ไม่ใช่คุณสมบัติหลักของแบคทีเรีย แบคทีเรียทั้งหมดจะถูกแสดงโดยเซลล์ชนิดพิเศษที่ไม่มีนิวเคลียสที่แท้จริงที่ล้อมรอบด้วยเยื่อหุ้มนิวเคลียส อะนาล็อกของนิวเคลียสในแบคทีเรียคือนิวเคลียส - พลาสมาที่มี DNA ซึ่งไม่ได้ถูกคั่นด้วยไซโตพลาสซึมด้วยเมมเบรน นอกจากนี้เซลล์แบคทีเรียยังมีลักษณะที่ไม่มีไมโตคอนเดรีย, คลอโรพลาสต์ตลอดจนโครงสร้างพิเศษและองค์ประกอบของโครงสร้างเมมเบรนและผนังเซลล์ สิ่งมีชีวิตที่เซลล์ขาดนิวเคลียสที่แท้จริงเรียกว่าโปรคาริโอต (พรีนิวเคลียร์) หรือโปรโตไซต์ (กล่าวคือ สิ่งมีชีวิตที่มีการจัดเรียงเซลล์ดั้งเดิม)[...]
เซลล์ Mycoplasma มีรูปร่างเป็นวงรีและมีขนาดเส้นผ่านศูนย์กลางประมาณ 0.1-0.25 นาโนเมตร (รูปที่ 43) มีลักษณะพิเศษคือการมีพลาสมาเมมเบรนชั้นนอกบาง ๆ (ความหนา - ประมาณ 8 นาโนเมตร) ซึ่งล้อมรอบไซโตพลาสซึมที่มีโมเลกุล DNA เพียงพอที่จะเข้ารหัสโปรตีนต่าง ๆ ประมาณ 800 ชนิด, RNA ประเภทต่าง ๆ, ไรโบโซมที่มีเส้นผ่านศูนย์กลางประมาณ 20 นาโนเมตร พลาสซึมของพวกมันประกอบด้วยสารต่างๆ มากมายในรูปของโปรตีน เม็ดไขมัน และสารประกอบอื่นๆ เนื่องจากเซลล์มีความแข็งแกร่งไม่เพียงพอ เยื่อมัยโคพลาสมาจึงผ่านตัวกรองแบคทีเรีย[...]
เป็นที่ยอมรับแล้วว่าการจับของกรดอะมิโนที่เปิดใช้งานเกิดขึ้นที่ไรโบโซมและการจัดวางลงในสายโซ่โพลีเปปไทด์ตามข้อมูลทางพันธุกรรมที่ได้รับจากนิวเคลียสผ่าน Messenger RNA (mRNA) ซึ่งจะอ่านข้อมูลที่เกี่ยวข้องจาก DNA และส่งไปยัง ไรโบโซม โปรตีนจำนวนหนึ่งถูกสังเคราะห์บนไรโบโซมที่แยกได้และมีการระบุการรวมกรดอะมิโนที่มีป้ายกำกับไว้ในนั้น บทบาทของเมทริกซ์ในการสังเคราะห์โปรตีนดำเนินการโดย mRNA ซึ่งติดอยู่กับไรโบโซม บนพื้นผิวของอย่างหลัง ปฏิสัมพันธ์เกิดขึ้นระหว่างกรดอะมิโนที่ซับซ้อน, การถ่ายโอน RNA ที่มีกรดอะมิโนตัวต่อไป, และลำดับนิวคลีโอไทด์ของ Messenger RNA ซึ่งทำหน้าที่บนไรโบโซมหนึ่งครั้งและหลังจากการสังเคราะห์สายโซ่โพลีเปปไทด์จะสลายตัว และโปรตีนที่สังเคราะห์ขึ้นใหม่จะสะสมอยู่ในไรโบโซม ในเซลล์แบคทีเรียที่มีระยะเวลาการงอกใหม่ 90 นาที อัตราการหมุนเวียนของ mRNA จะสูงถึง 4-6 วินาที[...]
ไซโตพลาสซึมเป็นสารละลายคอลลอยด์ ซึ่งมีระยะกระจายตัวเป็นสารประกอบโปรตีนเชิงซ้อนและสารที่คล้ายกับไขมัน และตัวกลางในการกระจายตัวคือน้ำ แบคทีเรียบางรูปแบบมีการรวมอยู่ในไซโตพลาสซึม - หยดของไขมัน, ซัลเฟอร์, ไกลโคเจน ฯลฯ ส่วนประกอบถาวรของเซลล์แบคทีเรียเป็นผลพลอยได้พิเศษของเยื่อหุ้มเซลล์ไซโตพลาสซึม - เมโซโซมซึ่งมีระบบรีดอกซ์ของเอนไซม์ ในการก่อตัวเหล่านี้ กระบวนการส่วนใหญ่เกี่ยวข้องกับการหายใจของแบคทีเรียเกิดขึ้น ในการรวมกันขนาดเล็ก - ไรโบโซมที่มีกรดไรโบนิวคลีอิกจะเกิดการสังเคราะห์โปรตีน แบคทีเรียส่วนใหญ่ไม่มีนิวเคลียสแยกจากกัน สารนิวเคลียร์ซึ่งแสดงโดย DNA ไม่ได้ถูกแยกออกจากไซโตพลาสซึมและก่อตัวเป็นนิวเคลียส การขนส่งสารที่จำเป็นสำหรับชีวิตของเซลล์และการกำจัดผลิตภัณฑ์เมตาบอลิซึมนั้นดำเนินการผ่านช่องทางและโพรงพิเศษซึ่งแยกออกจากไซโตพลาสซึมด้วยเมมเบรนที่มีโครงสร้างเหมือนกับไซโตพลาสซึม การก่อตัวของโครงสร้างนี้เรียกว่าเอนโดพลาสมิกเรติคูลัม (เรติคูลัม)[...]
ความคิดเกี่ยวกับความแปรปรวนและการถ่ายทอดทางพันธุกรรมของแบคทีเรียไม่สามารถเกิดขึ้นได้หากปราศจากความรู้เกี่ยวกับข้อกำหนดบางประการเกี่ยวกับอณูพันธุศาสตร์ของเซลล์โปรคาริโอต กระบวนการปรับตัวของการเพาะเลี้ยงจุลินทรีย์ให้เข้ากับสภาพแวดล้อมที่เปลี่ยนแปลงนั้นขึ้นอยู่กับความแปรปรวนและพันธุกรรม ซึ่งเป็นส่วนหนึ่งของพันธุกรรมของแบคทีเรีย เมื่อนำเสนอเซลล์วิทยาของเซลล์แบคทีเรีย โครงสร้างของ DNA และ RNA และบทบาทในชีวิตของเซลล์ได้รับการพิจารณาแล้ว โครงสร้างลักษณะเฉพาะของ DNA จะถูกรักษาไว้ในแต่ละสายพันธุ์และส่งต่อไปยังลูกหลานจากรุ่นสู่รุ่น เช่นเดียวกับลักษณะอื่นๆ DNA ของแบคทีเรียเป็นเกลียวสองเกลียวที่ปิดเป็นวงแหวน DNA ของแบคทีเรียที่มีวงแหวนซึ่งอยู่ในนิวเคลียสนั้นไม่มีโปรตีน วงแหวน DNA นี้สอดคล้องกับโครโมโซมของเซลล์ยูคาริโอต เป็นที่ทราบกันว่าโครโมโซมของเซลล์ยูคาริโอตนอกเหนือจาก DNA แล้วยังมีส่วนประกอบของโปรตีนอยู่เสมอ ตามมาว่าแนวคิดเรื่องโครโมโซมในยูคาริโอตค่อนข้างแตกต่างจากแนวคิดเรื่องโครโมโซมของแบคทีเรีย แน่นอนว่าสาย DNA ที่ประกอบเป็นโครโมโซมของแบคทีเรียนั้นแตกต่างกันไปในแต่ละสายพันธุ์ ส่วนประกอบของน้ำตาลฟอสเฟตของ DNA นั้นเหมือนกันในแบคทีเรียทุกประเภท ในทางกลับกันการจัดเรียงฐานไนโตรเจนและการรวมกันของพวกมันจะแตกต่างกันในแต่ละสายพันธุ์[...]
การใช้ยาปฏิชีวนะตามอำเภอใจที่เพิ่มขึ้นในปศุสัตว์ ซึ่งใช้ในปริมาณต่ำเพื่อส่งเสริมการเจริญเติบโตและยังเป็นมาตรการป้องกันความผิดปกติของระบบทางเดินอาหารที่เกี่ยวข้องกับความเครียดในสัตว์เลี้ยงในฟาร์ม กำลังนำไปสู่ความชุกของการดื้อยาปฏิชีวนะ R-factor ที่เพิ่มมากขึ้นในประชากรจุลินทรีย์ ส่งจากเซลล์แบคทีเรียหนึ่งไปยังอีกเซลล์หนึ่งในระหว่างการผันคำกริยา การถ่ายโอนเกิดขึ้นผ่านพลาสมิดซึ่งเป็น DNA ที่อยู่นอกโครโมโซมทรงกลมที่สามารถทำซ้ำได้[...]
ตรงกันข้ามกับฟาจที่มีความรุนแรง ที่เรียกว่าฟาจที่ออกฤทธิ์ปานกลาง หรือเรียกง่ายๆ ว่าฟาจปานกลาง ตัวแทนทั่วไปของฟาจดังกล่าวคือฟาจ X ซึ่งเคยเป็นและกำลังถูกใช้เป็นแบบจำลองการทดลองเพื่อชี้แจงประเด็นต่างๆ มากมายเกี่ยวกับพันธุศาสตร์ระดับโมเลกุล Phage X มีคุณสมบัติที่สำคัญสองประการ เช่นเดียวกับฟาจที่มีความรุนแรง มันสามารถแพร่เชื้อไปยังเซลล์แบคทีเรีย สืบพันธุ์แบบพืช สร้างสำเนาหลายร้อยสำเนาในเซลล์ และเซลล์ไลส์ปล่อยอนุภาคฟาจที่เจริญเต็มที่ อย่างไรก็ตาม DNA ของฟาจนี้สามารถรวมอยู่ในโครโมโซมของแบคทีเรียและกลายเป็นคำทำนายได้ ในกรณีนี้จะเกิดสิ่งที่เรียกว่าไลโซเจไนเซชันของแบคทีเรีย และแบคทีเรียที่มีการพยากรณ์โรคเรียกว่าไลโซเจนิก เซลล์แบคทีเรีย Lysogenic สามารถมีการพยากรณ์โรคได้เป็นเวลานานโดยไม่มีการสลาย การสลายด้วยการปล่อยอนุภาคฟาจใหม่จะถูกสังเกตหลังจากการสัมผัสกับแบคทีเรียไลโซเจนิกกับปัจจัยใดๆ เช่น รังสียูวี ซึ่งกระตุ้นให้เกิดการพัฒนาการพยากรณ์ให้เป็นฟาจ การศึกษาแบคทีเรียไลโซเจนิกทำให้สามารถรับข้อมูลใหม่จำนวนหนึ่งเกี่ยวกับบทบาทของโปรตีนต่างๆ ในการทำงานของยีนฟาจ[...]
จีโนมคลอโรพลาสต์ของพืชชั้นสูงจำนวนหนึ่งประกอบด้วย 120 ยีน จีโนมคลอโรพลาสต์มีความคล้ายคลึงกับจีโนมของแบคทีเรียมากทั้งในองค์กรและการทำงาน จีโนมไมโตคอนเดรียของมนุษย์อาจขาดอินตรอน แต่อินตรอนพบได้ใน DNA ของคลอโรพลาสต์ของพืชชั้นสูงบางชนิด เช่นเดียวกับใน DNA ของไมโตคอนเดรียของเชื้อรา เชื่อกันว่าจีโนมของคลอโรพลาสต์ของพืชชั้นสูงยังคงไม่เปลี่ยนแปลงเป็นเวลาประมาณหลายล้านปี เป็นไปได้ว่าสมัยโบราณดังกล่าวเป็นลักษณะเฉพาะของจีโนมไมโตคอนเดรียของสัตว์เลี้ยงลูกด้วยนม รวมถึงมนุษย์ด้วย[...]
ไดอะแกรมสมัยใหม่ที่แสดงการทำงานของยีนถูกสร้างขึ้นบนพื้นฐานของการวิเคราะห์เชิงตรรกะของข้อมูลการทดลองที่ได้รับโดยใช้วิธีทางชีวเคมีและพันธุกรรม การใช้กล้องจุลทรรศน์อิเล็กตรอนแบบละเอียดช่วยให้มองเห็นการทำงานของอุปกรณ์ทางพันธุกรรมของเซลล์ได้อย่างแท้จริง เมื่อเร็ว ๆ นี้ได้รับภาพถ่ายด้วยกล้องจุลทรรศน์อิเล็กตรอนซึ่งแสดงให้เห็นว่าเมทริกซ์ DNA ของแบคทีเรียในพื้นที่ที่มีโมเลกุลของ RNA polymerase (เอนไซม์ที่เร่งการถอดรหัสของ DNA เป็น RNA) ติดอยู่กับ DNA การสังเคราะห์โมเลกุล mRNA เกิดขึ้นได้อย่างไร . สาย mRNA ซึ่งตั้งฉากกับโมเลกุล DNA เชิงเส้นจะเคลื่อนที่ไปตามเมทริกซ์และเพิ่มความยาว เมื่อสาย RNA ยาวขึ้น ไรโบโซมก็จะเกาะติดกับมัน ซึ่งในทางกลับกัน เมื่อสาย RNA เคลื่อนตัวเข้าหา DNA จะนำไปสู่การสังเคราะห์โปรตีน[...]
การถ่ายโอนคือการถ่ายโอนสารพันธุกรรมจากแบคทีเรียผู้บริจาคไปยังแบคทีเรียผู้รับโดยใช้ฟาจ ปรากฏการณ์การทรานส์ดักชันถูกค้นพบครั้งแรกในปี พ.ศ. 2494 โดยเลเดอร์เบิร์กและเพื่อนร่วมงานของเขาในเชื้อ Salmonella typhimurium ปัจจุบันมีการแยกแยะความแตกต่างระหว่างการถ่ายทอดแบบไม่เฉพาะเจาะจงและแบบเฉพาะเจาะจง ด้วยการทรานส์ดักชันแบบไม่จำเพาะ ฟาจสามารถถ่ายโอนลักษณะใดๆ จากแบคทีเรียผู้ให้ไปยังแบคทีเรียผู้รับได้ การถ่ายโอนจะดำเนินการโดยฟาจระดับอุณหภูมิ (ไม่รุนแรง) เท่านั้น ฟาจอุณหภูมิปานกลางสามารถติดเชื้อแบคทีเรียได้ แต่อย่าแพร่พันธุ์ในพวกมันและไม่ทำให้เกิดการสลาย แต่จะรวมอยู่ใน DNA ของเซลล์แบคทีเรียและในสถานะที่ไม่ติดเชื้อในรูปแบบของการพยากรณ์ที่เรียกว่าจะถูกส่งจากเซลล์ ไปยังเซลล์ระหว่างการสืบพันธุ์ การเพาะเลี้ยงแบคทีเรียที่มีคำทำนายเรียกว่าไลโซเจนิก ในวัฒนธรรมเหล่านี้ ด้วยความถี่ต่ำ (ในหนึ่งใน 102 - 105 เซลล์) การสืบพันธุ์ของฟาจจะเกิดขึ้นเองและการสลายของเซลล์เกิดขึ้นพร้อมกับการปล่อยอนุภาคของฟาจ ซึ่งตรวจพบด้วยความช่วยเหลือของแบคทีเรียบ่งชี้ซึ่งฟาจดังกล่าวมีความรุนแรง .[...]
การทดลองดำเนินการในเซลล์สามห้อง ซึ่งประกอบด้วยห้องทำงานส่วนกลางและห้องอิเล็กโทรดสองห้อง วางสำลี 750 มก. ไว้ในห้องทำงานขนาด 25 X 7 X 37 มม. (ยาว X กว้าง X สูง) แยกออกจากห้องอิเล็กโทรดด้วยเยื่อกระดาษแก้ว โดยวิธีนี้ สารละลายเริ่มต้นของสารที่อยู่ระหว่างการศึกษาจะถูกป้อนจากล่างขึ้นบนด้วยความเร็วคงที่ เนื้อหาของสารประกอบในสารละลายเริ่มต้นที่จ่ายให้กับห้องทำงาน (C0) และในสารละลายที่ออกจากห้อง (Ci) ได้รับการตรวจสอบโดยการดูดซึมสูงสุดของโปรตีนและกรดนิวคลีอิกในช่วงหมายเลขคลื่น (35.5-38) X 103 ซม.- 1 โดยใช้เครื่องสเปกโตรโฟโตมิเตอร์ UV Specord UV-VIS ห้องอิเล็กโทรดเต็มไปด้วยถ่านกัมมันต์แบบเม็ด และน้ำกลั่นถูกส่งผ่านโดยแยกการไหล
สัณฐานวิทยาของแบคทีเรีย
แบคทีเรียจุลชีพมักเป็นสิ่งมีชีวิตเซลล์เดียวที่มีลักษณะเป็นพืช (จุลินทรีย์) แบคทีเรียบางชนิดมีลักษณะทางสัณฐานวิทยาที่มีความคงตัวเพียงพอ แบคทีเรียมีสามรูปแบบหลัก ได้แก่ ทรงกลมหรือวงรี (cocci) แท่ง (บาซิลลัส) และเกลียว
ค็อกซี่แบ่งออกเป็นคู่ - นักการทูต(เนสเซเรีย); tetracocci จัดเรียงเป็นกลุ่มละ 4 รูปเป็นรูปสี่เหลี่ยม cocci ที่ขึ้นรูปเป็นแพ็คเก็ตหรือ สารซาซินตั้งอยู่บน "พื้น"; สเตรปโตคอคกี้, เรียงกันเป็นโซ่; สตาฟิโลคอคกี้ก่อตัวเป็นกระจุกไร้รูปร่าง ชวนให้นึกถึงพวงองุ่น
แท่ง- ในบรรดาแท่งไม้ก็มี เดี่ยว, แบคทีเรียจัดเรียงแบบสุ่ม (enterobacteriaceae), ไดโพลบาซิลลัส,ตั้งอยู่เป็นคู่ (ตามบรรทัดเดียว) และ สเตรปโตบาซิลลัส,ก่อตัวเป็นโซ่ (แบคทีเรียแอนแทรกซ์)
แบคทีเรียที่มีรูปร่างเป็นเกลียวแบ่งออกเป็นสองกลุ่ม - วิบริโอและแบคทีเรียที่มีรูปร่างคล้าย ๆ กัน โดยส่วนโค้งของร่างกายไม่เกินหนึ่งในสี่ของเกลียว (Campylobacter) และ สไปโรเชเตสและสไปริลลามีส่วนโค้งเท่ากับเกลียวหนึ่งหรือหลายรอบ (สาเหตุของโรคซิฟิลิส)
แบคทีเรียใดๆ ประกอบด้วยองค์ประกอบ 3 ส่วน ได้แก่ โครงสร้างพื้นผิว เยื่อหุ้มเซลล์ ไซโตพลาสซึม
โครงสร้างพื้นผิวของแบคทีเรีย ได้แก่ แคปซูล แฟลเจลลา และไมโครวิลลี่
แคปซูลล้อมรอบเยื่อหุ้มเซลล์ของแบคทีเรียหลายชนิดรวมถึงแบคทีเรียที่ทำให้เกิดโรคด้วย แคปซูลขาดลักษณะการจัดองค์กรที่ได้รับคำสั่งของผนังเซลล์แบคทีเรีย มีไมโครแคปซูลซึ่งตรวจพบโดยกล้องจุลทรรศน์อิเล็กตรอนเท่านั้นในรูปแบบของชั้นของไมโครไฟบริล mucopolysaccharide) และแมคโครแคปซูล (ตรวจพบด้วยกล้องจุลทรรศน์แบบใช้แสง)
แคปซูลแบคทีเรียส่วนใหญ่ประกอบด้วยโพลีแซ็กคาไรด์เชิงซ้อน ตรวจพบได้โดยการย้อมสีตาม Burri-Gins หรือใช้ปฏิกิริยาบวมของ Neufeld แคปซูลอาจรวมถึงสารประกอบที่มีไนโตรเจน เช่น ของนิวโมคอกคัส (ประกอบด้วยโพลีแซ็กคาไรด์ กลูโคซามีน และกรดกลูโคโรนิก) แต่อาจไม่ประกอบด้วยไนโตรเจน เช่น แคปซูลลิวโคนอสตอค (ประกอบด้วยเดกซ์ทริน เลวูแลน ฟรุกโตซาน และโมโนแซ็กคาไรด์โพลีเมอร์อื่นๆ)
แคปซูลของแบคทีเรียบางชนิด (Bacillus anthracis) ประกอบด้วยโพลีแซ็กคาไรด์และโพลีเปปไทด์ที่เกิดจากโมโนเมอร์ของกรด D-กลูตามิก ซึ่งช่วยปกป้องแบคทีเรียจากเอนไซม์โปรตีโอไลติกของฟาโกไซต์
แฟลเจลลามีอยู่ในแบคทีเรียหลายชนิดและให้ความคล่องตัว แฟลเจลลัมเป็นเส้นใยโค้งเกลียวที่ขับเคลื่อนให้หมุนโดย "มอเตอร์" ซึ่งอยู่ที่จุดที่ยึดติดกับเมมเบรน ในแบคทีเรียที่แตกต่างกัน ความหนาของแฟลเจลลาจะแตกต่างกันไปตั้งแต่ 12 ถึง 18 นาโนเมตร และความยาวอาจสูงถึง 20 µm
แฟลเจลลาของแบคทีเรียประกอบด้วยโปรตีน (แฟลเจลลิน) และถูกสร้างขึ้นจากหน่วยย่อยที่มีน้ำหนักโมเลกุลค่อนข้างต่ำ เส้นใยของแฟลเจลลาถูกขับเคลื่อนด้วยตะขอฐานคล้ายบานพับเมมเบรน ซึ่งยึดไว้ด้วยตัวฐาน ซึ่งประกอบด้วยวงแหวนหนึ่งคู่ในแบคทีเรียแกรมบวก และวงแหวนสองคู่ในแบคทีเรียแกรมลบ วงแหวนทำหน้าที่เป็น "ดิสก์ไดรฟ์" และ "แบริ่ง" บนพื้นผิวด้านในของชั้นเพปทิโดไกลแคน โครงสร้างทั้งหมดทำหน้าที่ของตัวแปลงเคมีกล (มอเตอร์แฟลเจลลิน)
ที่ตั้ง.
อันตราย Flagella ตั้งอยู่บนพื้นผิวทั้งหมดของผนังเซลล์ (แบคทีเรียในตระกูล Enterobacteriaceae และ Bacillaceae)
โมโนทริชแฟลเจลลัมหนาหนึ่งอันที่ปลายด้านหนึ่ง (vibrios)
โพลิทริชแฟลเจลลา 2-50 มัด มองเห็นเป็นช่อเดียว
โพลาร์แฟลเจลลาติดอยู่ที่ปลายด้านใดด้านหนึ่งหรือทั้งสองด้านของแบคทีเรีย โลโฟทริช- มัดแฟลเจลลาที่ปลายด้านหนึ่งของแบคทีเรีย (ซูโดโมแนส) แอมฟิทริชชี่– มัดที่มีขั้วสองขั้ว (Spirillum)
ไมโครวิลลี่(pili, fimbriae) คือขนโปรตีน (ตั้งแต่ 10 ถึงหลายพัน) มีความหนา 3-25 นาโนเมตรและยาวสูงสุด 12 ไมครอน
ก. ดื่มธรรมดา.แบคทีเรียแกรมลบจำนวนมากมีพิลี (fimbriae) บางยาวที่เริ่มต้นจากเยื่อหุ้มเซลล์และทะลุผ่านผนังเซลล์ พวกมันถูกสร้างขึ้นโดยโปรตีนประเภทเดียวกันซึ่งมีโมเลกุลที่ก่อตัวเป็นเกลียว ของพวกเขา หน้าที่หลักคือการเกาะติดของแบคทีเรียกับสารตั้งต้นเช่นพื้นผิวเยื่อเมือกซึ่งเป็นปัจจัยสำคัญในการตั้งอาณานิคมและการติดเชื้อ นอกจากนี้การเพิ่มพื้นที่ผิวของเซลล์แบคทีเรียยังช่วยเพิ่มข้อได้เปรียบในการใช้สารอาหารจากสิ่งแวดล้อมอีกด้วย
บี. ฉ-ดื่ม(ปัจจัยการเจริญพันธุ์) – การก่อตัวพิเศษที่เกี่ยวข้องกับการผันของแบคทีเรีย มีลักษณะคล้ายหลอดโปรตีนกลวง ยาว 0.5-10 ไมครอน การก่อตัวของพวกมันถูกเข้ารหัสโดยพลาสมิด
เยื่อหุ้มเซลล์แบคทีเรียส่วนใหญ่ประกอบด้วยผนังเซลล์และเยื่อหุ้มเซลล์ไซโตพลาสซึม
ผนังเซลล์ของแบคทีเรียบาง ยืดหยุ่น และแข็ง และอาจไม่มีอยู่ในแบคทีเรียบางชนิดเลย (เช่น รูปแบบ L และไมโคพลาสมา) ผนังเซลล์ช่วยปกป้องแบคทีเรียจากอิทธิพลภายนอก ทำให้พวกมันมีรูปร่างที่มีลักษณะเฉพาะ และขนส่งสารอาหารและปล่อยสารเมตาบอไลต์ผ่านมัน บนพื้นผิวของมันมีตัวรับแบคทีเรียแบคทีเรียแบคทีเรียและสารเคมีต่างๆมากมาย CS รักษาความคงที่ของสภาพแวดล้อมภายในและทนทานต่อแรงกดดันที่สำคัญจากภายใน (เช่น ความดันบางส่วนของสารภายในเซลล์ของแบคทีเรียแกรมบวกสามารถสูงถึง 30 บรรยากาศ) โครงสร้างและองค์ประกอบขององค์ประกอบของ CS กำหนดความสามารถในการรับรู้สีย้อมเช่น ของพวกเขา คุณสมบัติของทิงทอเรียล- หลักการพื้นฐานประการหนึ่งในการสร้างความแตกต่างของแบคทีเรียคือความสามารถในการรับรู้และรักษาสีที่ซับซ้อนของเจนเชียนไวโอเล็ตที่มีไอโอดีนภายในเซลล์ หรือสูญเสียไปหลังการบำบัดด้วยแอลกอฮอล์ (แกรมสเตน) ดังนั้นจึงมีความแตกต่างระหว่างแกรมบวก (สีม่วงม่วง) และแกรมลบ (สีแดง)
ส่วนประกอบหลักของแบคทีเรีย CS คือ peptidoglycan (murein) Peptidoglycan มีค่อนข้างมากในแบคทีเรียแกรมบวก: สัดส่วนของโครงข่ายมูรินซึ่งมีความหนาประมาณ 40 ชั้น คิดเป็น 30–70% ของมวลแห้งของ CM แบคทีเรียแกรมลบประกอบด้วยมูรินเพียง 1-2 ชั้น ซึ่งคิดเป็นประมาณ 10% ของมวลแห้งของ CS
Peptidoglycan ถูกแสดงโดยโมเลกุลโพลีเมอร์ซึ่งประกอบด้วยกลุ่มไดแซ็กคาไรด์ซ้ำซึ่งการก่อตัวของมันเกี่ยวข้อง เอ็น-อะซิติลกลูโคซามีน และเอ็น-กรดอะซิติลมูรามิกหลังจับไดแซ็กคาไรด์ด้วยโอลิโกเปปไทด์ (จากกรดอะมิโนที่รู้จัก 20 ตัวใน CS ของแบคทีเรียพบเพียง 4 ตัวเท่านั้น - กรดกลูตามิก, ไกลซีน, ไลซีนและอะลานีน) แบคทีเรีย CS ยังรวมถึงกรดอะมิโนที่มีลักษณะเฉพาะ เช่น ไดมิโนพิเมลิกและ D-ไอโซเมอร์ของกรดกลูตามิกและอะลานีน ไลโซไซม์ไฮโดรไลซ์เปปทิโดไกลแคนโดยการตัดพันธะไกลโคซิดิกระหว่าง N-acetylglucosamine และกรด N-acetylmuramic
การเชื่อมโยงข้ามของเปปไทโดไกลแคนเกี่ยวข้องกับการก่อตัวของพันธะเปปไทด์ระหว่างเรซิดิวส่วนปลายของสายด้านข้างของเพปไทด์ (โดยปกติคือ D-อะลานีน) กับเรซิดิวสุดท้ายของสายด้านข้างที่อยู่ติดกัน (L-ไลซีนหรือกรดไดมิโนพิเมลิก)
แบคทีเรียแกรมบวกมี CS ที่มีการจัดระเบียบที่เรียบง่ายแต่ทรงพลัง ซึ่งประกอบด้วย peptidoglycan หลายชั้นเป็นส่วนใหญ่ รวมถึงลักษณะเฉพาะด้วย โพลีเมอร์กรดเทโชอิก– สายโซ่ของกลีเซอรอลหรือไรบิทอล 8-50 ตกค้าง เชื่อมต่อกันด้วยสะพานฟอสเฟต
แบคทีเรียแกรมลบมี CS ทินเนอร์ (เมื่อเทียบกับแบคทีเรียแกรมบวก) ซึ่งรวมถึงชั้น peptidoglycan สองโมเลกุลและไม่มีกรดเตอิโคอิก
ด้านบนของชั้นเพปทิโดไกลแคนจะมีเมมเบรนเพิ่มเติมหรือด้านนอก ความหนาเกินขนาดของชั้นเดียวของ peptidoglycan
ส่วนประกอบของเยื่อหุ้มชั้นนอก: ฟอสโฟไลปิด ไบเลเยอร์ โปรตีน โพลีแซ็กคาไรด์ และ LPS เรียงกันเป็นลวดลายโมเสก
ฟอสโฟไลปิด ไบเลเยอร์ยึดติดกับเพปทิโดไกลแคนโดยไลโปโปรตีนที่ข้ามช่องเพอริพลาสซึม
กระรอก, รวมทั้ง รูพรุนที่สร้างช่องทางของเมมเบรนเกี่ยวข้องกับการขนส่งไอออนและสารประกอบที่ชอบน้ำจากสภาพแวดล้อมภายนอกไปยังเพริพลาสซึม
แอลพีเอสสร้างขึ้นจากส่วนของไขมัน (ลิพิด A) แกนที่อุดมด้วยโพลีแซ็กคาไรด์ และโซ่ด้านข้างโพลีแซ็กคาไรด์ ส่วนโพลีแซ็กคาไรด์ของ LPS มีคุณสมบัติสร้างภูมิคุ้มกันและเรียกว่า O-Ag ส่วนไขมันมีความคงตัวต่อความร้อนและรับผิดชอบต่อผลกระทบทางชีวภาพของเอนโดทอกซิน
ออโตไลซิน- CS ของแบคทีเรียประกอบด้วยออโตไลซินซึ่งเป็นเอนไซม์ที่ละลายชั้นเปปทิโดไกลแคน กิจกรรมของพวกเขาจำเป็นสำหรับกระบวนการเจริญเติบโตของเซลล์ การแบ่งเซลล์ การสร้างสปอร์ และการบรรลุสภาวะของความสามารถในระหว่างการเปลี่ยนแปลง
เยื่อหุ้มเซลล์ไซโตพลาสซึม(หรือเมมเบรนของเซลล์หรือพลาสมา) เป็นสิ่งกีดขวางทางกายภาพ ออสโมติก และเมตาบอลิซึมระหว่างเนื้อหาภายในของเซลล์แบคทีเรียและสภาพแวดล้อมภายนอก CPM มีโครงสร้างสามชั้นที่ซับซ้อนและโดดเด่นด้วยความสามารถในการซึมผ่านแบบเลือกสรรที่เด่นชัด ในแบคทีเรียบางชนิดระหว่าง CPM และ CS จะมีช่องว่าง periplasmic - ช่องที่เต็มไปด้วยเอนไซม์ (ribonucleases, phosphatases, penicillinases ฯลฯ ) ในแบคทีเรียแกรมลบเอนไซม์จะถูกเทออกสู่สิ่งแวดล้อมอย่างอิสระ CPM ของแบคทีเรียประกอบด้วยโปรตีน ไขมัน คาร์โบไฮเดรต และ RNA
กระรอก CPM แบ่งออกเป็น โครงสร้างและ การทำงาน.อย่างหลังประกอบด้วยเอนไซม์ที่เกี่ยวข้องกับปฏิกิริยาสังเคราะห์บนพื้นผิวเมมเบรน กระบวนการรีดอกซ์ และเอนไซม์พิเศษบางชนิด (เช่น ซึมผ่าน).
ศูนย์กลางตั้งอยู่ ระบบขนส่งอิเล็กตรอนของแบคทีเรีย,ให้ความต้องการพลังงาน
เมโซโซม –การบุกรุกที่ซับซ้อนของ CPM ซึ่งยังไม่ได้รับการจัดตั้งขึ้นอย่างสมบูรณ์ เป็นที่รู้กันว่าเกี่ยวข้องกับนิวครอยด์และเกี่ยวข้องกับการแบ่งเซลล์และการสร้างสปอร์
การกำจัด CS ซึ่งป้องกัน CPM ที่อยู่ติดกัน นำไปสู่การสลายของแบคทีเรียหรือการก่อตัวของโปรโตพลาสต์และสฟีโรพลาสต์ ซึ่งมีต้นกำเนิดที่แตกต่างกัน (จากแบคทีเรียแกรมบวกหรือแกรมลบ ตามลำดับ) รวมถึงความเสถียรของการดูดซึม เมื่ออยู่ในสภาพแวดล้อมแบบไอโซโทนิก แบคทีเรียที่ขาด CS จะสามารถดูดซับ O 2 และปล่อย CO 2 ได้ รวมถึงเพิ่มจำนวนด้วย
ล-แบบฟอร์มภายใต้อิทธิพลของปัจจัยภายนอกบางประการ แบคทีเรียสามารถสูญเสีย CS ก่อตัวเป็นรูปตัว L (ตั้งชื่อตามสถาบัน D. Lister ซึ่งเป็นที่ที่พวกมันถูกแยกออกครั้งแรก) การเปลี่ยนแปลงดังกล่าวอาจเกิดขึ้นเอง (เช่นในหนองในเทียม) หรือเกิดขึ้น (เช่นภายใต้อิทธิพลของยาปฏิชีวนะ) ไฮไลท์ มั่นคงและไม่มั่นคงล-แบบฟอร์มแบบแรกไม่สามารถพลิกกลับได้ ในขณะที่แบบหลังกลับคืนสู่รูปแบบดั้งเดิมหลังจากกำจัดปัจจัยเชิงสาเหตุออกไปแล้ว
ตัวแทนของกลุ่มมัยโคพลาสมา (คลาส Mollicutes) ไม่มีผนังเซลล์
ไซโตพลาสซึมแบคทีเรีย - เมทริกซ์สำหรับการดำเนินการของปฏิกิริยาที่สำคัญ - ถูกแยกออกจาก CS ด้วยเยื่อหุ้มเซลล์ไซโตพลาสซึม ไซโตพลาสซึมของแบคทีเรียส่วนใหญ่ประกอบด้วย DNA, ไรโบโซม และเม็ดกักเก็บ พื้นที่ที่เหลือถูกครอบครองโดยเฟสคอลลอยด์ส่วนประกอบหลักของมันคือเอนไซม์ที่ละลายน้ำได้และ RNA (เมทริกซ์และการถ่ายโอน RNA) แบคทีเรียขาดคุณสมบัติออร์แกเนลต่างๆ ของเซลล์ยูคาริโอต และการทำงานของพวกมันจะดำเนินการโดย CPM ของแบคทีเรีย
ดีเอ็นเอ- เซลล์แบคทีเรียไม่มีเยื่อหุ้มนิวเคลียส DNA กระจุกตัวอยู่ในไซโตพลาสซึมในรูปของขดลวดที่เรียกว่านิวคลอยด์หรือจีโนฟอร์
เจโนฟอร์แบคทีเรียถูกแสดงโดยโมเลกุล DNA supercoiled ที่ปิดด้วยโควาเลนต์แบบเกลียวคู่ซึ่งคิดเป็น 2-3% ของมวลแห้งของเซลล์ (มากกว่า 10% โดยปริมาตร) ความยาวของรูปร่างโมเลกุลแตกต่างกันไปตั้งแต่ 0.25 ถึง 3 มม. DNA superhelix ของแบคทีเรียไม่มีฮิสโตน ปริมาณข้อมูลทางพันธุกรรมที่เข้ารหัสในยีนจะแตกต่างกันไปตามสปีชีส์ (ตัวอย่างเช่น จีโนม Escherichia coli เข้ารหัสโพลีเปปไทด์ที่แตกต่างกันประมาณ 4,000 ชนิด)
พลาสมิด- ในแบคทีเรีย อาจมีโมเลกุล DNA เพิ่มเติมอยู่ในรูปแบบขององค์ประกอบนอกโครโมโซมหรือรวมเข้ากับจีโนฟอร์ การรวมดังกล่าวเรียกว่าพลาสมิด (ตามลำดับ episomal หรือบูรณาการ- Episomal DNA นั้นมีลักษณะเป็นวงแหวนเช่นกัน แต่ episome นั้นมีขนาดเล็กกว่าโครโมโซมของแบคทีเรีย พลาสมิดมียีนที่แตกต่างกันจำนวนหนึ่งและมักจะระบุความรุนแรงของแบคทีเรีย แต่ข้อมูลที่อยู่ในพลาสมิดไม่จำเป็นอย่างยิ่งสำหรับเซลล์แบคทีเรีย
ไรโบโซมแบคทีเรียเป็นรูปแบบทรงกลมที่ซับซ้อนซึ่งประกอบด้วยโมเลกุล RNA ต่างๆ และโปรตีนที่เกี่ยวข้องหลายชนิด การก่อตัวทั้งหมดทำหน้าที่เป็นตำแหน่งของการสังเคราะห์โปรตีน
70 สไรโบโซม- เส้นผ่านศูนย์กลางของไรโบโซมของแบคทีเรียประมาณ 20 นาโนเมตร ค่าสัมประสิทธิ์การตกตะกอน – 70S (หน่วย Svedberg) ไรโบโซมของแบคทีเรียประกอบด้วยสองหน่วยย่อยที่มีค่าสัมประสิทธิ์การตกตะกอน 50S สำหรับหน่วยหนึ่งและ 30S สำหรับอีกหน่วย การรวมหน่วยย่อยเกิดขึ้นก่อนที่จะเริ่มการสังเคราะห์โปรตีน เซลล์แบคทีเรียสามารถมีไรโบโซมได้ตั้งแต่ 5,000 ถึง 50,000 ไรโบโซม ขึ้นอยู่กับความเข้มของการเจริญเติบโต
ยาปฏิชีวนะแบบแบคทีเรีย (สเตรปโตมัยซิน, เตตราไซคลิน, คลอแรมเฟนิคอล) ยับยั้งการสังเคราะห์โปรตีน, ปิดกั้นกระบวนการเผาผลาญบางอย่างที่เกิดขึ้นในไรโบโซมของแบคทีเรีย
เม็ดสำรองมีสารเมตาบอไลต์ส่วนเกินชั่วคราว การมีอยู่และจำนวนเม็ดจะแตกต่างกันไปขึ้นอยู่กับชนิดของแบคทีเรียและกิจกรรมการเผาผลาญของพวกมัน โพลีแซ็กคาไรด์ (แป้ง ไกลโคเจน แกรนูโลซา) ไขมัน (ไตรกลีเซอไรด์คล้ายกับไขมันของสัตว์ชั้นสูงถูกเก็บไว้ในยีสต์สกุล Candida ขี้ผึ้ง - ในมัยโคแบคทีเรียและโนคาร์เดีย โพลีเมอร์ของกรด β-ไฮดรอกซีบิวทีริก - เช่นในเซลล์ของบาซิลลัส megaterium) สามารถจัดเก็บได้ในรูปของเม็ด โพลีฟอสเฟต (เช่น โวลูติน พบครั้งแรกในสาหร่ายเกลียวทอง volutans) ซัลเฟอร์ (ในแบคทีเรียที่ออกซิไดซ์ซัลไฟด์เป็นซัลเฟต) โปรตีน - เช่น โปรทอกซิน (ใน Bacillus thuringiensis และสายพันธุ์ที่เกี่ยวข้อง) ).
ความลับหลักของสิ่งมีชีวิตอินทรีย์อยู่ที่ความสามารถในการทำซ้ำและส่งข้อมูลทางพันธุกรรมจากรุ่นก่อน ๆ ไปยังลูกหลานผ่านกลไกที่ค่อนข้างง่ายในการคัดลอกโมเลกุลขนาดใหญ่ของ DNA ของแต่ละเซลล์ที่มีชีวิตด้วยตนเอง แต่ละคนไม่ว่าสิ่งมีชีวิตจะประกอบด้วยเซลล์จำนวนมากหรือว่าเรากำลังพูดถึง DNA ที่พบในเซลล์ของแบคทีเรียก็ตามสิ่งมีชีวิตเซลล์เดียวที่เรียบง่ายเหล่านี้ซึ่งไม่สามารถรวบรวมได้เสมอไปแม้จะอยู่ในอาณานิคมขนาดใหญ่ก็ตาม .
เช่นเดียวกับตัวแทนของชีวิตอินทรีย์ ข้อมูลทางพันธุกรรม (ทางพันธุกรรม) ของแบคทีเรียจะถูกเก็บไว้ใน DNA ของพวกมัน ข้อมูลทางพันธุกรรมคืออะไร? โครงสร้างใดจัดเก็บข้อมูลทางพันธุกรรม
- ข้อมูลทางพันธุกรรมเป็นลำดับเฉพาะของนิวคลีโอไทด์ ไม่มีความลับอื่นใดในเคอร์เนล โดยการคัดลอกลำดับนี้ เซลล์จะสังเคราะห์โปรตีนหลากหลายชนิด พวกเขายังแก้ปัญหาอื่นๆ ทั้งหมดของร่างกาย ตั้งแต่ปัญหาขององค์กรไปจนถึงการจัดหาวัสดุก่อสร้างให้กับเซลล์
- โมเลกุลขนาดใหญ่ของ DNA ประกอบด้วยเบสนิวคลีอิกสี่เบส (อะดีนีน กัวนีน ไทมีน และไซโตซีน) ซึ่งรวมกันเป็นเกลียวคู่ด้วยน้ำตาลดีออกซีไรโบสและกรดฟอสฟอริกที่ตกค้าง เป็นฐานนิวคลีอิกที่เข้ารหัสลำดับการประกอบโปรตีน ไม่ว่าจะมีนิวเคลียสที่ก่อตัวอยู่ในเซลล์หรือไม่ก็ตาม
กรดดีออกซีไรโบนิวคลีอิกของแบคทีเรียมีโครงสร้างเหมือนกับโมเลกุลที่เก็บข้อมูลทางพันธุกรรมของสิ่งมีชีวิตอื่น ๆ ทั้งหมดบนโลก เช่นเดียวกับเซลล์อินทรีย์อื่นๆ แบคทีเรียจะสร้างโครโมโซมจากดีเอ็นเอ แต่นี่ไม่ได้หมายความว่าไม่มีความแตกต่างอื่น ๆ
ความแตกต่างพื้นฐานระหว่างแบคทีเรียคือไม่มีนิวเคลียสของเซลล์ ข้อมูลทางพันธุกรรมของแบคทีเรียไม่ได้ถูกรวบรวมไว้ในนิวเคลียสของเซลล์ มันเป็นเพียงโมเลกุลวงแหวนที่ติดอยู่กับผนังด้านหนึ่งของเยื่อหุ้มเซลล์ไซโตพลาสซึม
 อย่างไรก็ตาม ความจริงที่ว่าไม่มีแกนกลางไม่ได้ป้องกันกระบวนการจำลองและการแปลที่ใช้งานอยู่โดยใช้ผู้เก็บข้อมูลทางพันธุกรรมนี้ เพื่อให้เข้าใจว่าข้อมูลถูกถ่ายโอนอย่างไร คุณจำเป็นต้องเข้าใจว่าโครโมโซม ยีน และนิวเคลียสของเซลล์คืออะไร
อย่างไรก็ตาม ความจริงที่ว่าไม่มีแกนกลางไม่ได้ป้องกันกระบวนการจำลองและการแปลที่ใช้งานอยู่โดยใช้ผู้เก็บข้อมูลทางพันธุกรรมนี้ เพื่อให้เข้าใจว่าข้อมูลถูกถ่ายโอนอย่างไร คุณจำเป็นต้องเข้าใจว่าโครโมโซม ยีน และนิวเคลียสของเซลล์คืออะไร
- ยีนเป็นส่วนหนึ่งของโมเลกุลขนาดใหญ่ซึ่งมีการเขียนลำดับนิวคลีโอไทด์ที่ช่วยให้สามารถประกอบโปรตีนประเภทใดประเภทหนึ่งได้ ไม่มีข้อมูลอื่นในยีน
- โครโมโซมคือการรวมกันของสาย DNA กับโปรตีนฮิสโตนที่สร้างโครงสร้างและทำให้มีรูปร่างที่แน่นอนก่อนที่เซลล์จะเริ่มแบ่งตัว ในช่วงที่ไม่เกิดการแบ่งตัว จะไม่มีโครโมโซมเช่นนี้ในเซลล์ (หรือในนิวเคลียส หากเรากำลังพูดถึงยูคาริโอตนิวเคลียร์)
- นิวเคลียสของเซลล์เป็นโครงสร้างเซลล์ที่มีข้อมูลทางพันธุกรรมซึ่งจัดโครงสร้างเป็นโครโมโซมเมื่อเซลล์เตรียมที่จะแบ่งตัว มันเริ่มกระบวนการแบ่งแยกเอง สิ่งสำคัญคือต้องจำไว้ว่าแบคทีเรียไม่มีนิวเคลียสของเซลล์
หากในเซลล์ยูคาริโอตในระหว่างการแบ่งตัว มีการใช้โครงสร้างที่แยกจากกัน ซึ่งสร้างขึ้นเป็นพิเศษเพื่อความสะดวกในการแบ่งตัว แบคทีเรียจะขยายตัวได้อย่างไรในสภาวะที่ความสับสนวุ่นวายที่ยังไม่ปรากฏรูปแบบและชัดเจนในกรณีที่ไม่มีนิวเคลียสของเซลล์?
กรดดีออกซีไรโบนิวคลีอิกของเซลล์แบคทีเรีย
แม้ว่าโมเลกุล DNA ของแบคทีเรียจะแสดงเป็นโครงสร้างทรงกลมที่ค่อนข้างใหญ่ซึ่งอยู่ตรงกลางเซลล์ แต่จริงๆ แล้วเป็นรูปแบบที่ค่อนข้างกะทัดรัดซึ่งอยู่เฉพาะในพื้นที่ที่จำกัดของไซโตพลาสซึม
 เนื่องจากไม่มีเมมเบรนนิวเคลียร์ที่จะแยกโมเลกุลขนาดใหญ่ของแบคทีเรียที่รวมตัวกันออกจากโครงสร้างเซลล์อื่น เครื่องมือทางพันธุกรรมของสิ่งมีชีวิตที่ปราศจากนิวเคลียร์จึงไม่สามารถเชื่อมโยงกับเครื่องมือทางพันธุกรรมของยูคาริโอตได้ ดังนั้นเครื่องมือทางพันธุกรรมของโปรคาริโอตจึงถูกเรียกว่านิวครอยด์
เนื่องจากไม่มีเมมเบรนนิวเคลียร์ที่จะแยกโมเลกุลขนาดใหญ่ของแบคทีเรียที่รวมตัวกันออกจากโครงสร้างเซลล์อื่น เครื่องมือทางพันธุกรรมของสิ่งมีชีวิตที่ปราศจากนิวเคลียร์จึงไม่สามารถเชื่อมโยงกับเครื่องมือทางพันธุกรรมของยูคาริโอตได้ ดังนั้นเครื่องมือทางพันธุกรรมของโปรคาริโอตจึงถูกเรียกว่านิวครอยด์
ลักษณะเฉพาะของนิวครอยด์:
- DNA ที่มียีนหลายพันยีน
- ยีนถูกจัดเรียงเป็นเส้นตรงและเรียกว่าโครโมโซม โครโมโซมของแบคทีเรียคือชุดของยีนเชิงเส้น
- โมเลกุลขนาดใหญ่ยังถูกพับด้วยโปรตีนที่คล้ายกับฮิสโตนยูคาริโอต
นิวเคลียสติดอยู่กับเมมเบรนไซโตพลาสซึม ณ จุดที่การจำลองเริ่มต้นและสิ้นสุด (การคัดลอกด้วยตนเอง)

มีการทดลองแล้วว่านิวเคลียสและโครโมโซมไม่เหมือนกัน การเพิ่มจำนวนโครโมโซม (ยีนเชิงเส้น) เป็นหลักฐานว่าแบคทีเรียมีการแบ่งตัวอย่างแข็งขัน นิวคลอยด์หนึ่งตัวสามารถประกอบด้วยโครโมโซมหนึ่งโครโมโซมหรือหลายสำเนาก็ได้ ดังนั้นในช่วงเวลาการแบ่งตัว Azotobacter จะจำลองโครโมโซมเป็น 20-25 โครโมโซม (สำเนาของนิวครอยด์)
กระบวนการคัดลอก
ในการออกแบบทางทฤษฎีที่พัฒนาโดยนักจุลชีววิทยาในช่วงหลายปีที่ผ่านมาเมื่อเป็นเรื่องยากมากหรือเป็นไปไม่ได้เลยที่จะศึกษากระบวนการทางโมเลกุลที่ซับซ้อนด้วยการทดลอง การคัดลอกกรดดีออกซีไรโบนิวคลีอิกสามารถทำได้สามวิธี:
- อนุรักษ์นิยมซึ่งเกลียวคู่พาเรนต์ไม่คลายตัวและเกลียวคู่ของลูกสาวถูกสร้างขึ้นจากวัสดุใหม่ทั้งหมด
- กระจายตัวโดยที่โมเลกุลขนาดใหญ่แตกออกเป็นชิ้น ๆ และโมเลกุลลูกจะเกิดขึ้นบนลำดับนิวคลีโอไทด์ของชิ้นส่วนเหล่านี้เช่นเดียวกับในแม่แบบ
- กึ่งอนุรักษ์นิยม ตามแบบจำลองนี้ เกลียวคู่จะคลายตัว และแต่ละเกลียวของเกลียวทำหน้าที่เป็นแม่แบบสำหรับ DNA ของลูกสาว สิ่งที่เรียกว่าลูกผสมของโมเลกุลขนาดใหญ่และสายโซ่ที่สร้างขึ้นจากส่วนประกอบใหม่เกิดขึ้น
เมื่อในปี 1957 มีการค้นพบวิธีในการติดตามกระบวนการที่เกิดขึ้นใน DNA ของแบคทีเรียในระหว่างการจำลองแบบ พบว่ากรดดีออกซีไรโบนิวคลีอิกทำซ้ำในลักษณะกึ่งอนุรักษ์ นั่นคือ ผ่านการคลี่คลายและการใช้บริเวณที่คลี่คลายเป็นแม่แบบสำหรับการสังเคราะห์ โมเลกุลขนาดใหญ่ใหม่

กระบวนการจำลอง DNA ของแบคทีเรียนั้นคล้ายคลึงกับการจำลอง DNA ของกลไกอินทรีย์อื่น ๆ มากมันเกิดขึ้นตามรูปแบบดังต่อไปนี้:
- DNA helicase จะคลายและแตกเกลียวคู่โดยการเคลื่อนที่ไปตามกระดูกสันหลังของน้ำตาล-ฟอสเฟตของกรดดีออกซีไรโบนิวคลีอิก
- เอนไซม์โพลีเมอเรสเร่งปฏิกิริยาการเติมเบสนิวคลีอิกเสริมให้กับชิ้นส่วนของกรดดีออกซีไรโบนิวคลีอิกสายเดี่ยว
หลังจากการจำลองแบบ ส่วนหลักทั้งหมดของเซลล์จะถูกทำซ้ำ ได้แก่ ออร์แกเนลล์ เยื่อหุ้มเซลล์ไซโตพลาสซึม ผนังเซลล์ และเซลล์แบคทีเรียจะแยกออกเป็นสองส่วน
ปัญหา
นอกเหนือจากความสนใจทางวิทยาศาสตร์ล้วนๆ ในการศึกษา DNA ของแบคทีเรียแล้ว กลไกของการจำลองและการส่งข้อมูลทางพันธุกรรมจากเซลล์หนึ่งไปยังอีกเซลล์หนึ่งก็มีความสำคัญในทางปฏิบัติเป็นพิเศษเช่นกัน
เป็นที่ทราบกันดีว่าแบคทีเรียจะปรับตัวอย่างรวดเร็วเมื่อสัมผัสกับยาปฏิชีวนะ และเริ่มผลิตโปรตีนแอนติบอดีบางชนิดที่ขัดขวางผลการทำลายของยาปฏิชีวนะในเซลล์แบคทีเรีย ในแบคทีเรียรุ่นต่อๆ ไป ความต้านทานต่อยาต้านแบคทีเรียกลุ่มเฉพาะนี้จะยังคงอยู่
 ยิ่งไปกว่านั้น โดยการถ่ายโอนยีนแนวนอน (ไม่ผ่านการแบ่งตัว แต่โดยการสัมผัสแบคทีเรียตัวหนึ่งกับอีกแบคทีเรียหนึ่ง) ข้อมูลทางพันธุกรรมดังกล่าวก็จะถูกถ่ายโอนเช่นกัน ทำให้แบคทีเรียสายพันธุ์ต้านทานต่อยาปฏิชีวนะเพิ่มมากขึ้น
ยิ่งไปกว่านั้น โดยการถ่ายโอนยีนแนวนอน (ไม่ผ่านการแบ่งตัว แต่โดยการสัมผัสแบคทีเรียตัวหนึ่งกับอีกแบคทีเรียหนึ่ง) ข้อมูลทางพันธุกรรมดังกล่าวก็จะถูกถ่ายโอนเช่นกัน ทำให้แบคทีเรียสายพันธุ์ต้านทานต่อยาปฏิชีวนะเพิ่มมากขึ้น
การศึกษาคุณสมบัติเหล่านี้ของแบคทีเรีย การกำหนดว่ายีนภายนอกรวมอยู่ในโครงสร้างทั่วไปของกรดดีออกซีไรโบนิวคลีอิกอย่างไร เป็นสิ่งที่จุลชีววิทยาสมัยใหม่เกี่ยวข้อง


