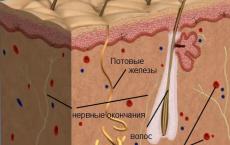
Каковы преимущества и недостатки явления гермафродитизма. Ну почему мы не гермафродиты? Возможные нарушения половой дифференцировки
Гермафродитизм (по имени греческого бога Гермафродита (др.-греч. Ἑρμαφρόδιτος )) - одновременное или последовательное наличие у организма мужских и женских половых признаков и репродуктивных органов.
Различают естественный гермафродитизм, присущий различным видам животных и растений (однодомность) и аномальный (патологический) гермафродитизм нормально раздельнополых животных (см. Гинандроморфизм, Интерсексуальность).
Естественный гермафродитизм
Гермафродитизм достаточно широко распространён в природе - как в растительном мире (в этом случае обычно используются термины однодомность или многодомность), так и среди животных. Большая часть высших растений являются гермафродитами, у животных гермафродитизм распространён прежде всего среди беспозвоночных - ряда кишечнополостных, подавляющего большинства плоских, некоторых кольчатых и круглых червей, моллюсков, ракообразных (в частности, большинства видов усоногих раков) и насекомых (кокциды).
Среди позвоночных гермафродитами являются многие виды рыб, причём наиболее часто гермафродитизм проявляется у рыб, населяющих коралловые рифы.
При естественном гермафродитизме особь способна продуцировать как мужские, так и женские гаметы, при этом возможна ситуация, когда способностью к оплодотворению обладают оба типа гамет (функциональный гермафродитизм), либо только один тип гамет (афункциональный гермафродитизм).
При синхронном гермафродитизме особь способна одновременно продуцировать и мужские, и женские гаметы.
В растительном мире такая ситуация зачастую приводит к самооплодотворению, встречающемуся у многих видов грибов, водорослей и цветковых растений (самоопыление у самофертильных растений).
В животном мире самооплодотворение при синхронном гермафродитизме встречается у гельминтов, гидр и моллюсков, а также некоторых рыб (Rivulus marmoratus ) однако в большинстве случаев автогамия предотвращается строением половых органов, при котором перенос собственных сперматозоидов в женские половые органы особи физически невозможен (моллюски, в частности, аплизии, ресничные черви), либо невозможностью слияния собственных дифференциированных гамет в жизнеспособную зиготу (некоторые асцидии).
Соответственно, при экзогамном синхронном гермафродитизме наблюдается два типа копулятивного поведения:
- взаимное оплодотворение, при котором обе копулирующие особи выполняют роль одновременно самцов и самок (наиболее часто среди беспозвоночных, в качестве примера можно привести дождевых червей, виноградных улиток)
- последовательное оплодотворение - одна из особей играет роль самца, а другая самки; взаимного оплодотворения в этом случае не происходит (например, у окуневых рыб родов Hypoplectrus и Serranus ).
Последовательный гермафродитизм (дихогамия)
В случае последовательного гермафродитизма (дихогамии) особь последовательно продуцирует мужские либо женские гаметы, при этом происходит либо последовательная активация гонад мужского и женского типов, либо смена фенотипа, ассоциированного с полом целиком. Дихогамия может проявляться как в пределах одного репродуктивного цикла, так и в течение жизненного цикла особи, при этом репродуктивный цикл может начинаться либо с мужской (протандрия), либо с женской (протогиния).
У растений, как правило, распространён первый вариант - при образовании цветков пыльники и рыльца созревают неодновременно. Таким образом, с одной стороны предотвращается самоопыление и, с другой стороны, за счёт неодновременности времени цветения различных растений в популяции, обеспечивается перекрёстное опыление.
В случае животных чаще всего происходит смена фенотипа, то есть смена пола. Ярким примером являются многие виды рыб - представителей семейств губановых (Labridae ), групперов (Serranidae ), помацентровых (Pomacentridae ), рыб-попугаев (Scaridae ), большинство которых являются обитателями коралловых рифов.
Аномальный (патологический) гермафродитизм
Наблюдается во всех группах животного мира, в том числе у высших позвоночных животных и человека. Гермафродитизм у людей является патологией сексуальной детерминации на генетическом или гормональном уровнях.
Различают истинный и ложный гермафродитизм:
- Истинный (гонадный) гермафродитизм характеризуется одновременным наличием мужских и женских половых органов, наряду с этим имеются одновременно мужские и женские половые железы. Яички и яичники при истинном гермафродитизме могут быть или объединены в одну смешанную половую железу, или располагаются отдельно. Вторичные половые признаки имеют элементы обоих полов: низкий тембр голоса, смешаный (бисексуальный) тип фигуры, в той или иной мере развитые молочные железы.
Хромосомный набор (кариотип) у таких больных обычно соответствует женскому кариотипу. В более редких случаях имеется ситуация, когда наличествуют как клетки, содержащие женский хромосомный набор, так и клетки, содержащие мужской хромосомный набор (явление так называемого мозаицизма). Истинный гермафродитизм - чрезвычайно редкое заболевание (в мировой литературе описано всего около 150 случаев) .
- Ложный гермафродитизм (псевдогермафродитизм) имеет место, когда налицо противоречие между внутренними (хромосомными и гонадными) и внешними (строение половых органов) признаками пола (бисексуальное развитие), то есть половые железы сформированы правильно по мужскому или женскому типу, но наружные половые органы имеют признаки двуполости.
С. Афонькин
Ну почему мы не гермафродиты?
Подыскивая подходящий подарок к 8 марта или с легким вздохом выполняя какойнибудь женский каприз, порой задумаешься над странным на первый взгляд вопросом: почему это матушка природа соизволила разделить род человеческий на отдельно существующие в пространстве половинки на женский и мужской пол? Мы так привыкли к этому делению, что воспринимаем его чуть ли не как единственно возможное.
Действительно, окружающие нас звери, птицы и рыбы, не говоря уже о большинстве других, более примитивных существ, тоже раздельнополы. Но некоторые демонстрируют нам совсем иное и вроде бы куда более разумное решение. И женская и мужская половые системы мирно уживаются у них в одном теле. Такое явление биологи назвали гермафродитизмом.
Легенда рассказывает, что Гермафродитом звали сына Гермеса и Афродиты, В прекрасного юношу влюбилась нимфа Салмакида, но тот не отвечал ей взаимностью. Обиженная невниманием нимфа подала жалобу в вышестоящую инстанцию самой Афродите, матери Гермафродита. А поскольку та занимала на Олимпе должность богини любви и была обязана потворствовать влюбленным, то недолго думая, со свойственной богам решимостью она слила нимфу и Гермафродита в единое существо чтобы последний не бегал от девушки.
Гермафродитизм у людей встречается чрезвычайно редко как патология на генетическом или гормональном уровнях. Среди животных, однако, попадаются виды, для которых гермафродитизм жизненная норма. Так устроены, например, некоторые ракообразные и многие черви.
Скептик, знакомый с биологией, может заметить, что возможное при гермафродитизме самооплодотворение чревато накоплением и проявлением генетических ошибок. Ведь одна из задач полового способа размножения хорошенько перемешать, перетасовать генетический материал родителей, что, в свою очередь, увеличивает разнообразие потомства и препятствует проявлению дефектов в генах. Так две книги с одним и тем же текстом, изданные в разных издательствах и в разные годы, вряд ли будут иметь одни и те же опечатки, и если читать их одновременно, то можно восстановить исходный текст без искажений. Точно такая же коррекция происходит и при совместной работе любой пары хромосом, одна из которых достается от отца, а другая от матери. Вспомните длительная практика политически выгодных династических браков между близкими родственниками нередко подрывала здоровье царствующих фамилий. Чего уж говорить о самооплодотворении самом близкородственном браке, какой можно вообразить!
Все это так. Однако у настоящих гермафродитов дело до самооплодотворения доходит редко. Чаще всего каждое такое существо выступает по очереди то в роли самца, то в роли самки. Если бы это было возможно у людей, отпали бы проблемы эмансипации, сексизма любой дискриминации, вызванной той или иной сексуальной ориентацией и принадлежностью к конкретному полу. Какая уж тут дискриминация при приеме на работу, если лучший мастер цеха Сидоров может в любое время уйти в декретный отпуск! Наверно, и семейные проблемы решались бы проще, если бы партнеры регулярно менялись ролями. А в случае разводов гермафродитных супружеских пар детей просто отдавали бы тому из супругов, который их родил. В крайних же обстоятельствах можно воспользоваться самооплодотворением. Например, Робинзон Крузо не грустил бы в одиночестве на необитаемом острове, а жил бы в кругу своей семьи.
Слов нет, преимущества гермафродитизма могут показаться соблазнительными и даже в чемто выгодными. Однако за редчайшими исключениями все позвоночные четко разделены на самцов и самок, а мы, люди, на мужчин и женщин. В чем же дело? Чем раздельнополость выгоднее и эволюционно привлекательнее гермафродитизма кроме уже упомянутой генетичекой выгоды?
Для того чтобы подвести под феномен разнополости теоретическую базу, доктор биологических наук Виген Артаваздович Геодакян создал собственную эволюционную теорию пола, которую давно и плодотворно развивает. Суть ее сводится к следующему.
Любая стремящаяся к самосохранению система (например, биологический вид), находящаяся в нестабильной, меняющейся со временем среде, пытается от этой среды обособиться и, в идеале, стать совершенно независимой от окружающих условий. Вся эволюция живого на Земле это длинный путь борьбы за независимость от прихотей окружающей среды. Путь пройден немалый от высыхающих во время отлива кембрийских медуз до современного человека с его кондиционерами и скафандрами.
Для того чтобы добиться столь впечатляющих успехов, живым существам постоянно приходилось решать две противоречащие друг другу задачи. С одной стороны, сохранять и передавать во времени уже сложившуюся информацию о собственном устройстве. С другой стороны, постоянно реагировать на ее изменения, подлаживаться под ее меняющиеся свойства, искать способы ей противодействовать и, в конечном счете, вводить соответствующие коррективы в передаваемую по цепочке поколений информацию. Ибо куда же от нее, среды, деться?
На заре эволюции, вероятно, еще во времена царствования одноклеточных, решение этих разных стратегических задач было возложено на два противоположных пола, потому что отдавать два противоречивых приказа одному подчиненному глупо и неэффективно. Женский пол взял на себя функцию сохранять и передавать, а мужской исследовать и привносить изменения в сохраняемое. Говоря образно, женский пол взял на себя консервативную роль, а мужской революционную.
До появления техники оценивать среду обитания можно было лишь одним способом ценой своей жизни. Подходишь среде обитания живи и плодись. Не подходишь извини... Для того, чтобы лучше выполнять свою миссию, мужской пол обзавелся целым рядом любопытных свойств. У мужских особей выше частота мутаций своеобразных случайных проб по принципу: "а что получится, если сделать так? ". У особей мужского пола четче и однозначнее проявляются все генетически определяемые признаки. У людей этот феномен приводит к тому, что однояйцевые близнецымальчики больше похожи друг на друга, чем близнецыдевочки.
С другой стороны, фенотипическое разнообразие представителей всего мужского пола в целом больше, чем женского опятьтаки, чтобы жизни было легче выбирать среди них. Например, частота ранней детской смертности выше у мальчиков, но зато и среди долгожителей мужчин больше, чем женщин. Среде было из чего выбрать.
Мужчины чаще лезут на рожон, у них выше поисковая активность и агрессивность. Случайность? Вовсе нет! Именно такое поведение гарантирует качественный "отстрел " средой плохо приспособленных индивидов. Средняя продолжительность жизни у мужчин меньше, чем у женщин. Случайность? Нет, результат страшного давления среды на непластичных мужчин! Реальный срок жизни мужчин почти всегда меньше отведенного им природой. Точно так же среднестатистический воробей живет на свободе меньше года, а в клетке спокойно чирикает два десятка лет. Какие воробьи доживут на свободе до второго года и обзаведутся, наконец, семьей? Стреляные, но недостреленные средой обитания.
Если же вода, огонь и медные трубы окружающей среды успешно пройдены, если среда поставила свой штамп ОТК на данном генотипе, у его носителя есть все шансы активно распространить свои гены в следующем поколении, став владельцем многочисленного гарема или на худой конец просто многодетным папашей. Вы не задумывались, почему природа создала такое неравноправие между полами в возможности оставлять потомство? Женский пол здесь ограничен. Сколько детей может родить женщина за всю жизнь? В лучшем случае десятка три? А падишах или популярный певец? Тут счет может пойти уже на тысячи, если не на десятки тысяч потомков.
Подобный перекос, только в соотношении половых клетокгамет, возник еще в мире одноклеточных. Еще в кембрийском океане одну царственно огромную женскую гамету водоросли или какойнибудь медузы окружала целая толпа мужских гамет. Для репродуктивного успеха каждому из этих кавалеров предстояло совершить поистине геракловы подвиги: потратить массу энергии для дальнего путешествия, проплыть огромное (с клеточной точки зрения) расстояние, найти по запаху желанную клетку и слиться с ней раньше других. По сути, ту же картину мы наблюдаем при оплодотворении у большинства современных существ.
Биологи говорят о гаметическом отборе, происходящем до оплодотворения. В результате внутриутробного марафона подавляющее большинство мужских сперматозоидов сходит с дистанции и просто не добирается до яйцеклетки. Проверяются ли на этом этапе их жизненные способности? Вполне вероятно. Говорят, что за счет эффекта гаметического отбора частота генетических нарушений в потомстве людей, переживших бомбардировку Хиросимы, оказалась гораздо ниже прогнозируемой.
Представительницы женского пола более пластичны. Это позволяет ловко уходить от давления среды, сохранять доставшийся набор генов независимо от того, что досталось. Другими словами, приспосабливаться к любой жизненной ситуации, явно или подсознательно думая только об одном о детях. О возможности и необходимости передать им палочку генетической эстафеты.
В стабильной среде разница между мужским и женским полом сводится к чисто анатомическим различиям репродуктивных органов. Иначе говоря, половой диморфизм сведен к минимуму. Однако стоит среде начать меняться, как сразу начинается расхождение полов по целому ряду признаков, которые могут иметь приспособительное значение. Вначале начинает меняться мужской пол ценой своей жизни он перебирает возможные варианты ответа на изменившиеся условия. Затем, после отбора наиболее приемлемых вариантов, следом за ним начинает подтягиваться "осторожный " женский пол. Другими словами, любой поразному выраженный у двух полов признак меняется во времени от женской его формы к мужской.
Взглянув на половой диморфизм под таким углом зрения, можно четко увидеть направление эволюционного изменения вида или целой группы видов. Например, у большинства позвоночных самцы крупнее самок. Следовательно, эволюция этой группы идет по пути постепенного увеличения средних размеров тела. Взгляните на график изменения среднего роста людей за последние несколько тысяч лет, и вы поймете, почему мужчины в среднем несколько выше женщин.
Футуристическая, устремленная в будущее природа изменений, свойственных мужскому полу, ярко проявляется в характере врожденных патологий. У женского пола эти аномалии носят атавистическую природу, то есть напоминают об уже пройденных этапах эволюции. У мужского же пола они направлены в будущее, то есть как бы продолжают вектор уже наметившихся изменений. Например, среди новорожденных с тремя почками вдвое больше девочек, чем мальчиков, и наоборот среди новорожденных однопочечников преобладают мальчики. Такая же картина и в случае аномального числа ребер. В полном соответствии с Ветхим Заветом мальчишки с меньшим их количеством рождаются чаще девчонок. Почему так? Все просто! Эволюция всех хордовых шла по пути уменьшения числа гомологичных органов (в том числе почек и ребер). Оба пола демонстрируют отголоски этого процесса поразному. Женский показывает что было, мужской что будет.
Гермафродитизм (по имени греческого бога Гермафродита греч.Ερμαφρόδιτος) - одновременное или последовательное наличие у организма мужских и женских половых признаков и репродуктивных органов.
Различают естественный гермафродитизм, присущий различным видам животных и растений (однодомность) и аномальный (патологический) гермафродитизм нормально раздельнополых животных (см. Гинандроморфизм,Интерсексуальность).
Гермафродитизм достаточно широко распространён в природе - как в растительном мире (в этом случае обычно используются термины однодомность или многодомность), так и среди животных. Большая часть высших растений являются гермафродитами, у животных гермафродитизм распространён прежде всего среди беспозвоночных - рядакишечнополостных, подавляющего большинства плоских, некоторых кольчатых и круглых червей, моллюсков,ракообразных (в частности, большинства видов усоногих раков) и насекомых (кокциды).
Среди позвоночных гермафродитами являются многие виды рыб, причём наиболее част гермафродитизм у рыб, населяющих коралловые рифы.
При естественном гермафродитизме особь способна продуцировать как мужские, так и женские гаметы, при этом возможна ситуация, когда способностью к оплодотворению обладают оба типа гамет (функциональный гермафродитизм), либо только один тип гамет (афункциональный гермафродитизм).
При синхронном гермафродитизме особь способна одновременно продуцировать и мужские, и женские гаметы.
В растительном мире такая ситуация зачастую приводит к самооплодотворению, встречающемуся у многих видов грибов, водорослей и цветковых растений (самоопыление у самофертильных растений).
В животном мире самооплодотворение при синхронном гермафродитизме встречается у гельминтов, гидр и моллюсков, а также некоторых рыб (Rivulus marmoratus) однако в большинстве случаев автогамия предотвращается строением половых органов, при котором перенос собственных сперматозоидов в женские половые органы особи физически невозможен (моллюски, в частности, аплизии, ресничные черви), либо невозможностью слияния собственных дифференциированных гамет в жизнеспособную зиготу (некоторые асцидии).
Соответственно, при экзогамном синхронном гермафродитизме наблюдается два типа копулятивного поведения:
взаимное оплодотворение, при котором обе копулирующие особи выполняют роль одновременно самцов и самок (наиболее часто среди беспозвоночных, в качестве примера можно привести дождевых червей, виноградных улиток)
последовательное оплодотворение - одна из особей играет роль самца, а другая самки; взаимного оплодотворения в этом случае не происходит (например, у окуневых рыб родов Hypoplectrus и Serranus).
В случае последовательного гермафродитизма (дихогамии) особь последовательно продуцирует мужские либо женские гаметы, при этом происходит либо последовательная активация гонад мужского и женского типов, либо смена фенотипа, ассоциированного с полом целиком. Дихогамия может проявляться как в пределах одного репродуктивного цикла, так и в течение жизненного цикла особи, при этом репродуктивный цикл может начинаться либо с мужской (протандрия), либо с женской (протогиния).
У растений, как правило, распространён первый вариант - при образовании цветков пыльники и рыльца созревают неодновременно. Таким образом, с одной стороны предотвращается самоопыление и, с другой стороны, за счёт неодновременности времени цветения различных растений в популяции, обеспечивается перекрёстное опыление.
В случае животных чаще всего происходит смена фенотипа, то есть смена пола. Ярким примером являются многие виды рыб - представителей семейств губановых (Labridae), групперов (Serranidae), помацентровых (Pomacentridae), рыб-попугаев (Scaridae), большинство которых являются обитателями коралловых рифов.
Патологический гермафродитизм наблюдается во всех группах животного мира, в том числе у высших позвоночных животных и человека. Гермафродитизм у людей является патологией сексуальной детерминации на генетическом или гормональном уровнях.
Различают истинный и ложный гермафродитизм:
Истинный (гонадный) гермафродитизм характеризуется одновременным наличием мужских и женских половых органов, наряду с этим имеются одновременно мужские и женские половые железы. Яички и яичники при истинном гермафродитизме могут быть или объединены в одну смешанную половую железу, или располагаются отдельно. Вторичные половые признаки имеют элементы обоих полов: низкий тембр голоса, смешаный (бисексуальный) тип фигуры, в той или иной мере развитые молочные железы.
Хромосомный набор (кариотип) у таких больных обычно соответствует женскому кариотипу. В более редких случаях имеется ситуация, когда наличествуют как клетки, содержащие женский хромосомный набор, так и клетки, содержащие мужской хромосомный набор (явление так называемого мозаицизма). Истинный гермафродитизм - чрезвычайно редкое заболевание (в мировой литературе описано всего около 150 случаев).
Ложный гермафродитизм (псевдогермафродитизм) имеет место, когда налицо противоречие между внутренними (хромосомными и гонадными) и внешними (строение половых органов) признаками пола (бисексуальное развитие), т.е.половые железы сформированы правильно по мужскому или женскому типу, но наружные половые органы имеют признаки двуполости.
Гинандроморфизм (др.-греч. γυνή - женщина + ἀνήρ, род. п. ἀνδρός - мужчина + μορφή - вид, форма) - аномалия, выражающаяся в том, что в одном организме крупные участки тела имеют генотип и признаки разных полов. Является результатом наличия в мужских и женских клетках организма наборовполовых хромосом с разным количеством последних, как например у многих насекомых. Гинандроморфизм происходит как результат неправильного распределения половых хромосом по клеткам в ходе нарушенного созревания яйцеклетки, её оплодотворения или дробления.
Особи - гинандроморфы наиболее ярко выражены у насекомых с четко проявляющимися признаками полового диморфизма, при этом морфологически выделяются следующие типы гинандроморфов:
билатеральные, у которых одна продольная половина тела имеет признаки мужского пола, другая - женского;
передне-задние, у которых передняя часть тела несет признаки одного пола, а задняя - другого;
мозаичные, у которых перемежаются участки тела, несущие признаки разных полов.
У позвоночных животных и у человека вследствие действия половых гормонов подобные явления приводят к половым аномалиям, при которых секториальное распределение мужских и женских тканей обычно проявляется не так резко.
При интерсексуальности наблюдается более сложная дифференциация женских и мужских признаков.
Интерсексуальность - наличие у раздельнополого организма признаков обоих полов, причем эти признаки являются не полностью развитыми, промежуточными (ср. Гермафродитизм). Признаки обоих полов проявляются совместно на одних и тех же частях тела (ср. Гинандроморфизм).
Эмбриональное развитие такого организма называется интерсексом, начинается нормально, но с определённого момента продолжается по типу другого пола. Чем раньше меняется направление развития организма, тем резче выражена у него интерсексуальность.
Является результатом отклонения от нормы набора половых хромосом и генов в момент оплодотворения при соединении гамет в зиготу. По характеру нарушения бывает триплоидная или иная - анеуплоидная интерсекуальность. Диплоидная интерсексуальность наблюдается при скрещивании разных географических рас у бабочки непарного шелкопряда, причем либо у самок, либо у самцов, в зависимости от типа скрещивания.
Формы интерсексуальности, так называемого псевдогермафродитизма у человека, также могут быть вызваны нарушением нормального числа половых хромосом. При этом у мух дрозофил определяющим в развитии пола является соотношение числа пар половых хромосом и аутосом, поэтому у них интерсексуальность обычно связана с нарушением этого соотношения (например, наблюдается при соотношении 3А:2X - три набора аутосом на две половые хромосомы). У человека определяющим фактором развития мужского пола является наличие У-хромосомы, при этом черты интерсексуальности наблюдаются у мужчин с синдромом Клайнфельтера(набор половых хромосом ХХУ).
Гормонная интерсексуальность. Если у животных выделения половыми железами мужских или женских гормонов определяют развитие вторичных половых признаков, то у них можно наблюдать явление гормональной интерсексуальности.
Билет 13
1. Провизорные органы, виды и формирование образований провизорных клеток
Провизорные органы (нем. provisorisch - предварительный, временный) - временные органы зародышей иличинок многоклеточных животных, функционирующие только в эмбриональный или личиночный период развития. Могут выполнять функции, специфические для зародыша или личинки, или основные функции организма до формирования аналогичных дефинитивных (окончательных) органов, свойственных для взрослого организма.
Примеры провизорных органов: хорион, амнион, желточный мешок, аллантоис и серозная оболочка и другие.
Амнион - временный орган, обеспечивающий водную среду для развития зародыша. В эмбриогенезе человека он появляется на второй стадии гаструляции сначала как небольшой пузырек, дном которого является первичная эктодерма (эпибласт) зародыша
Амниотическая оболочка образует стенку резервуара, заполненного амниотической жидкостью, в которой находится плод.
Основная функция амниотической оболочки - выработка околоплодных вод, обеспечивающих среду для развивающегося организма и предохраняющих его от механического повреждения. Эпителий амниона, обращенный в его полость, не только выделяет околоплодные воды, но и принимает участие в обратном всасывании их. В амниотической жидкости поддерживаются до конца беременности необходимый состав и концентрация солей. Амнион выполняет также защитную функцию, предупреждая попадание в плод вредоносных агентов.
Желточный мешок -орган, депонирующий питательные вещества (желток), необходимые для развития зародыша. У человека он образован внезародышевой энтодермой и внезародышевой мезодермой (мезенхимой). Желточный мешок является первым органом, в стенке которого развиваются кровяные островки, формирующие первые клетки крови и первые кровеносные сосуды, обеспечивающие у плода перенос кислорода и питательных веществ.
Аллантоис - небольшой отросток в отделе зародыша, врастающий в амниотическую ножку. Он является производным желточного мешка и состоит из внезародышевой энтодермы и висцерального листка мезодермы. У человека аллантоис не достигает значительного развития, но его роль в обеспечении питания и дыхания зародыша все же велика, так как по нему к хориону растут сосуды, располагающиеся в пупочном канатике.
Пупочный канатик - представляет собой упругий тяж, соединяющий зародыш (плод) с плацентой.
Дальнейшее развития хориона связано с двумя процессами - разрушением слизистой оболочки матки вследствие протеолитической активности наружного слоя и развитием плаценты.
Плацента (детское место) человека относится к типу дискоидальных гемохориальных ворсинчатых плацент. Плацента обеспечивает связь плода с материнским организмом, создает барьер между кровью матери и плода.
Функции плаценты: дыхательная; транспорт питательных веществ, воды, электролитов; выделительная; эндокринная; участие в сокращении миометрия.
Самооплодотворяющиеся животные размножаются при прочих равных вдвое быстрее, чем раздельнополые. Почему же раздельнополость преобладает в природе? Для ответа на этот вопрос были искусственно выведены породы круглых червей Caenorhabditis elegans , одни из которых практикуют только перекрестное оплодотворение, другие — только самооплодотворение. Эксперименты с этими червями подтвердили две гипотезы о преимуществах перекрестного оплодотворения. Одно преимущество состоит в более эффективном очищении генофонда от вредных мутаций, второе — в ускоренном накоплении полезных мутаций, что помогает популяции приспосабливаться к меняющимся условиям.
|
|
Двойная цена самцов
Зачем нужно половое размножение, зачем нужны самцы? Ответы на эти вопросы совсем не так очевидны, как может показаться.
|

Известный эволюционист Джон Мэйнард Смит (John Maynard Smith) обратил внимание на серьезность данной проблемы в своей книге The Evolution of Sex (1978). Мэйнард Смит подробно рассмотрел парадокс, которому дал название «двойная цена пола» (two-fold cost of sex). Суть его в том, что при прочих равных бесполое размножение (или самооплодотворение) ровно в два раза эффективнее, чем перекрестное оплодотворение с участием самцов (см. рисунок). Иными словами, самцы обходятся популяции непомерно дорого. Отказ от них дает немедленный и очень значительный выигрыш в скорости размножения. Мы знаем, что чисто технически переход от раздельнополости и перекрестного оплодотворения к бесполому размножению или самооплодотворению вполне возможен, тому есть масса примеров как у растений, так и у животных (см., например: Самки гигантского комодского варана размножаются без участия самцов, «Элементы», 26.12.2006). Тем не менее бесполые расы и популяции самооплодотворяющихся гермафродитов почему-то до сих пор так и не вытеснили тех, кто размножается «обычным» образом, с участием самцов.
Зачем они все-таки нужны?
Из сказанного следует, что перекрестное оплодотворение должно давать некие преимущества, настолько значительные, что они перекрывают даже двойной выигрыш в эффективности размножения, даваемый отказом от самцов. Причем эти преимущества должны проявляться сразу, а не когда-нибудь через миллион лет. Естественному отбору нет дела до отдаленных перспектив.
Есть много гипотез о природе этих преимуществ (см.: Evolution of sexual reproduction). Мы рассмотрим две из них. Первая известна под названием «храповик Мюллера» (см.: Muller"s ratchet). Храповик — это устройство, в котором ось может крутиться только в одну сторону. Суть идеи в том, что если у бесполого организма возникает вредная мутация, его потомки уже не могут от нее избавиться. Она будет, как родовое проклятие, передаваться всем его потомкам вечно (если только не произойдет обратная мутация, а вероятность этого очень мала). У бесполых организмов отбор может отбраковывать только целые геномы, но не отдельные гены. Поэтому в череде поколений бесполых организмов может (при соблюдении определенных условий) происходить неуклонное накопление вредных мутаций. Одним из таких условий является достаточно большой размер генома. У круглых червей, между прочим, геномы маленькие по сравнению с другими животными. Может быть, поэтому они и могут позволить себе самооплодотворение (см. ниже).
Если же организмы размножаются половым путем и практикуют перекрестное оплодотворение, то индивидуальные геномы постоянно рассыпаются и перемешиваются, а новые геномы формируются из фрагментов, ранее принадлежавших разным организмам. В результате возникает особая новая сущность, которой нет у бесполых организмов, — генофонд популяции. Гены получают возможность размножаться или выбраковываться независимо друг от друга. Ген с неудачной мутацией может быть отбракован отбором, а остальные («хорошие») гены данного родительского организма могут благополучно сохраниться в популяции.
Таким образом, первая идея состоит в том, что половое размножение способствует очищению геномов от «генетического груза», то есть помогает избавляться от постоянно возникающих вредных мутаций, не допуская вырождения (снижения общей приспособленности популяции).
Вторая идея сродни первой: она предполагает, что половое размножение помогает организмам эффективнее адаптироваться к меняющимся условиям за счет ускоренного накопления мутаций, полезных в данной обстановке. Допустим, у одной особи возникла одна полезная мутация, у другой — другая. Если эти организмы бесполые, у них практически нет шансов дождаться объединения обеих мутаций в одном геноме. Половое размножение дает такую возможность. Оно фактически делает все полезные мутации, возникшие в популяции, «общим достоянием». Ясно, что скорость приспособления к меняющимся условиям у организмов с половым размножением должна быть выше.
Все эти теоретические построения, однако, основаны на определенных допущениях. Результаты математического моделирования свидетельствуют о том, что степень полезности или вредности перекрестного оплодотворения по сравнению с бесполым размножением или самооплодотворением зависит от целого ряда параметров. В их числе размер популяции; скорость мутирования; размер генома; количественное распределение мутаций в зависимости от степени их вредности/полезности; число потомков, производимых одной самкой; эффективность отбора (степень зависимости числа оставляемых потомков не от случайных, а от генетических факторов) и т. д. Некоторые из этих параметров очень трудно измерить не только в природных, но и в лабораторных популяциях.
Поэтому все гипотезы такого рода остро нуждаются не столько в теоретических обоснованиях и математических моделях (всего этого уже в избытке), сколько в прямой экспериментальной проверке. Однако подобных экспериментов до сих пор проведено не так уж много (Colegrave, 2002. Sex releases the speed limit on evolution // Nature . V. 420. P. 664-666; Goddard et al., 2005. Sex increases the efficacy of natural selection in experimental yeast populations // Nature . V. 434. P. 636-640). Новое исследование, выполненное биологами из Орегонского университета на круглом черве Caenorhabditis elegans , ярко проиллюстрировало действенность обоих рассмотренных механизмов, обеспечивающих преимущество тем популяциям, которые не отказываются от самцов, несмотря на их «двойную цену».
Уникальный объект для изучения роли самцов
Черви Caenorhabditis elegans словно нарочно созданы для экспериментальной проверки вышеупомянутых гипотез. У этих червей нет самок. Популяции состоят из самцов и гермафродитов, причем последние численно преобладают. У гермафродитов две X-хромосомы, у самцов — только одна (система определения пола X0, как у дрозофилы). Гермафродиты производят сперматозоиды и яйцеклетки и могут размножаться без посторонней помощи путем самооплодотворения. Самцы производят только сперматозоиды и могут оплодотворять гермафродитов. В результате самооплодотворения на свет появляются только гермафродиты. При перекрестном оплодотворении половина потомства оказывается гермафродитами, половина — самцами. Обычно частота перекрестного оплодотворения в популяциях C. elegans не превышает нескольких процентов. Чтобы определить эту частоту, не обязательно наблюдать за интимной жизнью червей — достаточно знать процент самцов в популяции.
Следует пояснить, что самооплодотворение — это не совсем то же самое, что бесполое (клональное) размножение, однако различия между ними быстро сходят на нет в череде самооплодотворяющихся поколений. Самооплодотворяющиеся организмы за несколько поколений становятся гомозиготными по всем локусам. После этого потомство перестает отличаться от родителей генетически, точно так же, как и при клональном размножении.
У C. elegans известны мутации, влияющие на частоту перекрестного оплодотворения. Одна из них, xol-1 , смертельна для самцов и фактически приводит к тому, что в популяции остаются только гермафродиты, размножающиеся путем самооплодотворения. Другая, fog-2 , лишает гермафродитов способности производить сперму и фактически превращает их в самок. Популяция, в которой все особи несут эту мутацию, становится обычной раздельнополой популяцией, как у большинства животных.
Авторы при помощи классических методик (путем скрещиваний, а не генной инженерии) вывели две пары пород червей с почти одинаковыми геномами, различающимися только наличием мутаций xol-1 и fog-2 . Первая порода в каждой паре, с мутацией xol-1, размножается только самооплодотворением (obligate selfing, OS). Вторая, с мутацией fog-2, может размножаться только путем перекрестного оплодотворения (obligate outcrossing, OO). К каждой паре пород прилагалась третья, с таким же генетическим «бэкграундом», но лишенная обеих мутаций (wild type, WT). У пород WT частота перекрестного оплодотворения в стандартных лабораторных условиях не превышает 5%.
Самцы нужны! Проверено экспериментально
С этими тройками пород были проведены две серии экспериментов.
В первой серии проверялась гипотеза о том, что перекрестное оплодотворение помогает избавляться от «генетического груза». Эксперимент продолжался в течение 50 поколений (червей, разумеется, а не экспериментаторов). Каждое поколение червей подвергалось действию химического мутагена — этилметансульфоната. Это приводило к увеличению частоты мутирования примерно в четыре раза. Молодых животных помещали в чашку Петри, разделенную пополам стенкой из вермикулита (см. рисунок), причем червей сажали в одну половину чашки, а их пища — бактерии E. coli — находилась в другой половине. Червей при пересадке обрабатывали антибиотиком, чтобы очистить от случайно прилипших бактерий. В результате для того, чтобы добраться до еды, а значит получить шанс выжить и оставить потомство, черви должны были преодолеть препятствие. Тем самым экспериментаторы повысили эффективность «очищающего» отбора, который отсеивает вредные мутации. В обычных лабораторных условиях эффективность отбора очень низка, потому что черви окружены пищей со всех сторон. В такой ситуации могут выжить и размножиться даже очень слабые, перегруженные вредными мутациями животные. В новой экспериментальной установке этой уравниловке был положен конец. Чтобы переползти через стенку, червь должен быть здоров и крепок.
Авторы сравнивали приспособленность (fitness) у червей до и после эксперимента, то есть у особей первого и пятидесятого поколения. Червей C. elegans можно долго хранить в замороженном виде. Это сильно облегчает подобные эксперименты. Пока длился эксперимент, выборка червей 1-го поколения спокойно лежала в морозильнике. Приспособленность измеряли следующим образом. Червей смешивали в равной пропорции с контрольными червями, в геном которых был вставлен ген светящегося белка, и сажали в экспериментальную установку. Животным давали время, чтобы преодолеть барьер и размножиться, а затем определяли процент несветящихся особей в потомстве. Если этот процент увеличился в пятидесятом поколении по сравнению с первым — значит, в ходе эксперимента приспособленность выросла, если уменьшился — значит, имело место вырождение.
Результаты эксперимента показаны на рисунке. Они однозначно свидетельствуют о том, что перекрестное оплодотворение является мощным средством борьбы с генетическим грузом. Чем выше частота перекрестного оплодотворения, тем лучше итоговый результат (все линии на рисунке возрастают слева направо). Искусственно повышенная скорость мутирования привела к вырождению (снижению приспособленности) всех пород червей, кроме OO — «облигатных перекрестников».
Даже тем породам, у которых мутагенез не был искусственно ускорен, высокая частота перекрестного оплодотворения дала преимущество. В обычных лабораторных условиях это преимущество не проявляется, потому что червям не нужно перелезать через стенки, чтобы добраться до корма.
Любопытно, что в одной из двух контрольных пород OS («облигатных самооплодотворителей») даже без повышения скорости мутирования отказ от перекрестного оплодотворения привел к вырождению (левый квадратик в верхней паре кривых на рисунке расположен ниже нуля).
Рисунок также показывает, что частота перекрестного оплодотворения у большинства «диких» пород (WT) в ходе эксперимента оказалась заметно выше исходных 5%. Это, пожалуй, самый важный результат. Он означает, что в жестких условиях (имеется в виду как необходимость перелезать через барьер, так и повышенный темп мутагенеза) естественный отбор дает явное преимущество особям, размножающимся путем перекрестного оплодотворения. Потомство таких особей оказывается более жизнеспособным, и поэтому в ходе эксперимента происходит отбор на склонность к перекрестному оплодотворению.
Таким образом, первый эксперимент убедительно подтвердил гипотезу о том, что перекрестное оплодотворение помогает популяции избавляться от вредных мутаций.
Во второй серии экспериментов проверялось, помогает ли перекрестное оплодотворение вырабатывать новые адаптации путем накопления полезных мутаций. На этот раз червям, чтобы добраться до пищи, нужно было преодолеть зону, заселенную патогенными бактериями Serratia . Эти бактерии, попадая в пищеварительный тракт C. elegans , вызывают у червя опасное заболевание, которое может кончится смертью. Чтобы выжить в этой ситуации, черви должны были либо научиться не глотать вредных бактерий, либо выработать устойчивость к ним. Какой из вариантов выбрали подопытные популяции червей — неизвестно, однако за 40 поколений породы OO отлично приспособились к новым условиям, породы WT приспособились несколько хуже, а породы OS не приспособились совсем (их выживаемость в среде с вредными бактериями осталась на исходном низком уровне). И снова в ходе эксперимента у пород WT под действием отбора резко возросла частота перекрестного оплодотворения.
Таким образом, перекрестное оплодотворение действительно помогает популяции приспосабливаться к меняющимся условиям, в данном случае — к появлению болезнетворного микроба. Тот факт, что в ходе эксперимента у пород WT увеличивалась частота перекрестного оплодотворения, означает, что спаривание с самцами (в противовес самооплодотворению) дает гермафродитам немедленное адаптивное преимущество, которое, очевидно, перевешивает «двойную цену», которую им приходится платить, производя на свет самцов.
Следует отметить, что перекрестное оплодотворение встречается не только у раздельнополых организмов. Например, многие беспозвоночные являются гермафродитами, оплодотворяющими не сами себя, а друг друга — перекрестно. У растений перекрестное опыление обоеполых («гермафродитных») особей тоже, мягко говоря, не редкость. Обе гипотезы, проверявшиеся в данной работе, вполне приложимы к таким гермафродитам. Иными словами, эта работа не доказала, что «перекрестный гермафродитизм» в чём-то уступает раздельнополости. А ведь за первый из этих двух вариантов не нужно платить пресловутую «двойную цену». Следовательно, проблема всё равно остается.
Проведенные эксперименты выявили недостатки самооплодотворения по сравнению с перекрестным, но они не объяснили, почему многие организмы предпочли раздельнополость «перекрестному гермафродитизму». Ключом к решению этой загадки, скорее всего, является половой отбор. Раздельнополость дает возможность самкам придирчиво выбирать себе партнеров, и это может служить дополнительным способом повышения эффективности отбраковки вредных и накопления полезных мутаций. Возможно, и эта гипотеза когда-нибудь получит экспериментальные подтверждения.
Шла речь о дождевых червях. Елена задала очень хороший вопрос о размножении червей, и затронула тему биологического гермафродитизма. За что ей отдельное и большое СПАСИБО, поскольку у меня совсем недавно поселились 6 очень маленьких и интересных гермафродита в баночке, и я выжидала момента, чтобы познакомить вас с ними.
Гермафродитизм, как биологическое явление, предполагает наличие у особи и женских и мужских гонад (половых желёз), производящих как женские, так и мужские гаметы (половые клетки). Женские гаметы называются яйцеклетки, а мужские – сперматозоиды. Размножение путем гермафродитизма относится к половому, поскольку в нем принимают участие гаметы.
Название явления восходит к греческой мифологии. Гермафродит – это сын богини любви Афродиты и
бога торговли, магии и разума Гермеса. Он был необычайно прекрасен. И однажды
он встретился с юной нимфой Салмакидой. Нимфа так страстно и безответно влюбилась
в Гермафродита, что попросила богов навеки соединить их. Боги исполнили ее мольбу
и пара слилась в одно существо.
Рисунок, кстати, сделан с помощью цикория)))
В мире животных гермафродитизм чрезвычайно популярное явление среди беспозвоночных. Так типы кишечнополостные, плоские, кольчатые черви, многие моллюски размножаются с помощью естественного гермафродитизма.
У высших позвоночных форм гермафродитизм в норме представлен у отдельных видов рыб. У амфибий, рептилий, птиц и млекопитающих (в том числе и у человека), гермафродитизм рассматривается как явление патологии (нарушения) эмбрионального (зародышевого) развития.
Кроме того различают синхронный и последовательный гермафродитизм.
При последовательном гермафродитизме. У особи созревает только один тип гамет. Например, у рыбы-попугая, сначала активируется женская половая система, продуцирующая икру, тогда рыба будет самкой. Через некоторое время, рыба меняет пол на мужской, и под действием гормонов производит сперматозоиды, то есть становится самцом.
При синхронном, одновременно продуцируются яйцеклетки и сперматозоиды в пределах особи. Чаще бывает перекрестный синхронный гермафродитизм, например, у пиявок и дождевых червей. Когда дождевые черви готовы к размножению, у них образуется муфточка на нескольких передних кольцевых сегментах, которая называется поясок.
В пояске находятся яйцеклетки. При встрече
два червя обмениваются сперматозоидами,
которые оплодотворяют яйцеклетки внутри муфты. Когда происходит слияние
яйцеклетки и сперматозоида, образуется зигота, или оплодотворенное яйцо. Всего
в муфточке может находиться до 20 яиц. На фотографии отчетливо видна муфточка.
Через некоторое время после оплодотворения, муфточка соскальзывает с червя. Ее стенки затвердевают, цвет с желтоватого меняется на коричневый, и формируется кокон. Из кокона выходят маленькие червячки около 1 мм в длину. Такие муфточки формируются у половозрелого червя каждую неделю.
Явление гермафродитизма повышает шансы особей на оплодотворение. Ведь минимальное количество особей для обмена генетическим материалом сводится до двух при любом раскладе. В случае раздельнополых существ, вероятность уже не 100%, а только 50%.
Но здесь возрастает коэффициент вариативности потомства, оно будет с более разнообразными признаками, а значит, получит больше шансов приспособиться к меняющимся условиям окружающей среды и выжить, поэтому у более сложных форм животных есть самцы и самки.
С месяц назад у меня появились очаровательные маленькие гермафродиты – 6 улиток ахатин. Накануне я мечтала, хотя бы об одной, а тут их появилось шесть. Мне их принесла и подарила моя коллега математик. Для нее было очень большим сюрпризом, что их улитки отложили кладку яиц, из которых вывелись малышата. Я не смогла их оставить в кабинете биологии, поскольку за малышами нужен ежедневный уход, забрала домой чтобы подрастить. Когда подрастут, раздам жаждущим.
Улитки кушают только салат латук, остальной салат,
огурцы, яблоки и капусту дружно игнорируют вшестером. Сегодня, наконец,
сфотографировала улиточек. Это ночные
существа, днем они спят, закапываясь в кокосовый грунт. Просыпаются в 10-11
часов вечера и начинают натирать листочек на своей радуле (терке-языке). Если
прислушаться, то даже слышно как они хрустят.
Даю им еще толченую скорлупу.
Сквозь прозрачные покровы можно увидеть, как продвигается проглоченный кусочек
скорлупки по пищеварительному тракту. Двигаются они размеренно, наблюдать за ними медитативное удовольствие. Сначала выложила улиточек в цепочку, но вскоре они устроили небольшую кучу-малу. Самая маленькая улиточка любит исследовать, она больше других передвигается по банке. А самая большая улитка любит поесть.